In the fields of molecular biology and genetics, a genome is all the genetic information of an organism. It consists of nucleotide sequences of DNA. The nuclear genome includes protein-coding genes and non-coding genes, other functional regions of the genome such as regulatory sequences, and often a substantial fraction of junk DNA with no evident function. Almost all eukaryotes have mitochondria and a small mitochondrial genome. Algae and plants also contain chloroplasts with a chloroplast genome.

A transposable element is a nucleic acid sequence in DNA that can change its position within a genome, sometimes creating or reversing mutations and altering the cell's genetic identity and genome size. Transposition often results in duplication of the same genetic material. In the human genome, L1 and Alu elements are two examples. Barbara McClintock's discovery of them earned her a Nobel Prize in 1983. Its importance in personalized medicine is becoming increasingly relevant, as well as gaining more attention in data analytics given the difficulty of analysis in very high dimensional spaces.

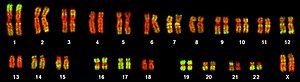
The human genome is a complete set of nucleic acid sequences for humans, encoded as DNA within the 23 chromosome pairs in cell nuclei and in a small DNA molecule found within individual mitochondria. These are usually treated separately as the nuclear genome and the mitochondrial genome. Human genomes include both protein-coding DNA sequences and various types of DNA that does not encode proteins. The latter is a diverse category that includes DNA coding for non-translated RNA, such as that for ribosomal RNA, transfer RNA, ribozymes, small nuclear RNAs, and several types of regulatory RNAs. It also includes promoters and their associated gene-regulatory elements, DNA playing structural and replicatory roles, such as scaffolding regions, telomeres, centromeres, and origins of replication, plus large numbers of transposable elements, inserted viral DNA, non-functional pseudogenes and simple, highly repetitive sequences. Introns make up a large percentage of non-coding DNA. Some of this non-coding DNA is non-functional junk DNA, such as pseudogenes, but there is no firm consensus on the total amount of junk DNA.
Non-coding DNA (ncDNA) sequences are components of an organism's DNA that do not encode protein sequences. Some non-coding DNA is transcribed into functional non-coding RNA molecules. Other functional regions of the non-coding DNA fraction include regulatory sequences that control gene expression; scaffold attachment regions; origins of DNA replication; centromeres; and telomeres. Some non-coding regions appear to be mostly nonfunctional, such as introns, pseudogenes, intergenic DNA, and fragments of transposons and viruses. Regions that are completely nonfunctional are called junk DNA.

Pseudogenes are nonfunctional segments of DNA that resemble functional genes. Most arise as superfluous copies of functional genes, either directly by gene duplication or indirectly by reverse transcription of an mRNA transcript. Pseudogenes are usually identified when genome sequence analysis finds gene-like sequences that lack regulatory sequences needed for transcription or translation, or whose coding sequences are obviously defective due to frameshifts or premature stop codons. Pseudogenes are a type of junk DNA.
Repeated sequences are short or long patterns of nucleic acids that occur in multiple copies throughout the genome. In many organisms, a significant fraction of the genomic DNA is repetitive, with over two-thirds of the sequence consisting of repetitive elements in humans. Some of these repeated sequences are necessary for maintaining important genome structures such as telomeres or centromeres.

Retrotransposons are a type of genetic component that copy and paste themselves into different genomic locations (transposon) by converting RNA back into DNA through the reverse transcription process using an RNA transposition intermediate.
Interspersed repetitive DNA is found in all eukaryotic genomes. They differ from tandem repeat DNA in that rather than the repeat sequences coming right after one another, they are dispersed throughout the genome and nonadjacent. The sequence that repeats can vary depending on the type of organism, and many other factors. Certain classes of interspersed repeat sequences propagate themselves by RNA mediated transposition; they have been called retrotransposons, and they constitute 25–40% of most mammalian genomes. Some types of interspersed repetitive DNA elements allow new genes to evolve by uncoupling similar DNA sequences from gene conversion during meiosis.

Endogenous retroviruses (ERVs) are endogenous viral elements in the genome that closely resemble and can be derived from retroviruses. They are abundant in the genomes of jawed vertebrates, and they comprise up to 5–8% of the human genome.
Retrotransposon markers are components of DNA which are used as cladistic markers. They assist in determining the common ancestry, or not, of related taxa. The "presence" of a given retrotransposon in related taxa suggests their orthologous integration, a derived condition acquired via a common ancestry, while the "absence" of particular elements indicates the plesiomorphic condition prior to integration in more distant taxa. The use of presence/absence analyses to reconstruct the systematic biology of mammals depends on the availability of retrotransposons that were actively integrating before the divergence of a particular species.
Exon shuffling is a molecular mechanism for the formation of new genes. It is a process through which two or more exons from different genes can be brought together ectopically, or the same exon can be duplicated, to create a new exon-intron structure. There are different mechanisms through which exon shuffling occurs: transposon mediated exon shuffling, crossover during sexual recombination of parental genomes and illegitimate recombination.

Mobile genetic elements (MGEs), sometimes called selfish genetic elements, are a type of genetic material that can move around within a genome, or that can be transferred from one species or replicon to another. MGEs are found in all organisms. In humans, approximately 50% of the genome is thought to be MGEs. MGEs play a distinct role in evolution. Gene duplication events can also happen through the mechanism of MGEs. MGEs can also cause mutations in protein coding regions, which alters the protein functions. These mechanisms can also rearrange genes in the host genome generating variation. These mechanism can increase fitness by gaining new or additional functions. An example of MGEs in evolutionary context are that virulence factors and antibiotic resistance genes of MGEs can be transported to share genetic code with neighboring bacteria. However, MGEs can also decrease fitness by introducing disease-causing alleles or mutations. The set of MGEs in an organism is called a mobilome, which is composed of a large number of plasmids, transposons and viruses.
In the fields of bioinformatics and computational biology, Genome survey sequences (GSS) are nucleotide sequences similar to expressed sequence tags (ESTs) that the only difference is that most of them are genomic in origin, rather than mRNA.
A conserved non-coding sequence (CNS) is a DNA sequence of noncoding DNA that is evolutionarily conserved. These sequences are of interest for their potential to regulate gene production.
An endogenous viral element (EVE) is a DNA sequence derived from a virus, and present within the germline of a non-viral organism. EVEs may be entire viral genomes (proviruses), or fragments of viral genomes. They arise when a viral DNA sequence becomes integrated into the genome of a germ cell that goes on to produce a viable organism. The newly established EVE can be inherited from one generation to the next as an allele in the host species, and may even reach fixation.

Long interspersed nuclear elements (LINEs) are a group of non-LTR retrotransposons that are widespread in the genome of many eukaryotes. LINEs contain an internal Pol II promoter to initiate transcription into mRNA, and encode one or two proteins, ORF1 and ORF2. The functional domains present within ORF1 vary greatly among LINEs, but often exhibit RNA/DNA binding activity. ORF2 is essential to successful retrotransposition, and encodes a protein with both reverse transcriptase and endonuclease activity.

LINE1 is a family of related class I transposable elements in the DNA of some organisms, classified with the long interspersed elements (LINEs). L1 transposons comprise approximately 17% of the human genome. These active L1s can interrupt the genome through insertions, deletions, rearrangements, and copy number variations. L1 activity has contributed to the instability and evolution of genomes and is tightly regulated in the germline by DNA methylation, histone modifications, and piRNA. L1s can further impact genome variation through mispairing and unequal crossing over during meiosis due to its repetitive DNA sequences.
Short interspersed nuclear elements (SINEs) are non-autonomous, non-coding transposable elements (TEs) that are about 100 to 700 base pairs in length. They are a class of retrotransposons, DNA elements that amplify themselves throughout eukaryotic genomes, often through RNA intermediates. SINEs compose about 13% of the mammalian genome.
DNA transposons are DNA sequences, sometimes referred to "jumping genes", that can move and integrate to different locations within the genome. They are class II transposable elements (TEs) that move through a DNA intermediate, as opposed to class I TEs, retrotransposons, that move through an RNA intermediate. DNA transposons can move in the DNA of an organism via a single-or double-stranded DNA intermediate. DNA transposons have been found in both prokaryotic and eukaryotic organisms. They can make up a significant portion of an organism's genome, particularly in eukaryotes. In prokaryotes, TE's can facilitate the horizontal transfer of antibiotic resistance or other genes associated with virulence. After replicating and propagating in a host, all transposon copies become inactivated and are lost unless the transposon passes to a genome by starting a new life cycle with horizontal transfer. It is important to note that DNA transposons do not randomly insert themselves into the genome, but rather show preference for specific sites.
Haig Hagop Kazazian Jr. was an American professor in the Department of Genetic Medicine at Johns Hopkins University School of Medicine in Baltimore, Maryland. Kazazian was an elected member of the National Academy of Sciences and the American Academy of Arts and Sciences.