
Ascomycota is a phylum of the kingdom Fungi that, together with the Basidiomycota, forms the subkingdom Dikarya. Its members are commonly known as the sac fungi or ascomycetes. It is the largest phylum of Fungi, with over 64,000 species. The defining feature of this fungal group is the "ascus", a microscopic sexual structure in which nonmotile spores, called ascospores, are formed. However, some species of Ascomycota are asexual and thus do not form asci or ascospores. Familiar examples of sac fungi include morels, truffles, brewers' and bakers' yeast, dead man's fingers, and cup fungi. The fungal symbionts in the majority of lichens such as Cladonia belong to the Ascomycota.

A hypha is a long, branching, filamentous structure of a fungus, oomycete, or actinobacterium. In most fungi, hyphae are the main mode of vegetative growth, and are collectively called a mycelium.
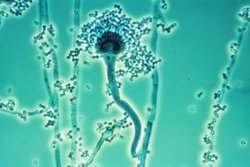
Aspergillus fumigatus is a species of fungus in the genus Aspergillus, and is one of the most common Aspergillus species to cause disease in individuals with an immunodeficiency.
This is a glossary of some of the terms used in phytopathology.
Hyphomycetes are a form classification of fungi, part of what has often been referred to as fungi imperfecti, Deuteromycota, or anamorphic fungi. Hyphomycetes lack closed fruit bodies, and are often referred to as moulds. Most hyphomycetes are now assigned to the Ascomycota, on the basis of genetic connections made by life-cycle studies or by phylogenetic analysis of DNA sequences; many remain unassigned phylogenetically.

Nematophagous fungi are carnivorous fungi specialized in trapping and digesting nematodes. Around 160 species are known. Species exist that live inside the nematodes from the beginning and others that catch them, mostly with glue traps or in rings, some of which constrict on contact. Some species possess both types of traps. Another technique is to stun the nematodes using toxins, a method employed by Coprinus comatus, Stropharia rugosoannulata, and the family Pleurotaceae. The habit of feeding on nematodes has arisen many times among fungi, as is demonstrated by the fact that nematophagous species are found in all major fungal groups. Nematophagous fungi can be useful in controlling those nematodes that eat crops. Purpureocillium, for example, can be used as a bio-nematicide.

The Orbiliaceae are a family of saprobic sac fungi. It is the only family in the monotypic class Orbiliomycetes and the monotypic order Orbiliales. The family was first described by John Axel Nannfeldt in 1932 and now contains 288 species in 12 genera. Members of this family have a widespread distribution, but are more prevalent in temperate regions. Some species in the Orbiliaceae are carnivorous fungi, and have evolved a number of specialized mechanisms to trap nematodes.

Purpureocillium lilacinum is a species of filamentous fungus in the family Ophiocordycipitaceae. It has been isolated from a wide range of habitats, including cultivated and uncultivated soils, forests, grassland, deserts, estuarine sediments and sewage sludge, and insects. It has also been found in nematode eggs, and occasionally from females of root-knot and cyst nematodes. In addition, it has frequently been detected in the rhizosphere of many crops. The species can grow at a wide range of temperatures – from 8 to 38 °C for a few isolates, with optimal growth in the range 26 to 30 °C. It also has a wide pH tolerance and can grow on a variety of substrates. P. lilacinum has shown promising results for use as a biocontrol agent to control the growth of destructive root-knot nematodes.

Zoophagus is a genus of zygomycete fungi that preys on rotifers and nematodes. It was established in 1911 by Sommerstorff, who originally considered it to be an oomycete. It is common in a variety of freshwater habitats, such as ponds and sewage treatment plants.

Trichothecium roseum is a fungus in the division Ascomycota first reported in 1809. It is characterized by its flat and granular colonies which are initially white and develop to be light pink in color. This fungus reproduces asexually through the formation of conidia with no known sexual state. Trichothecium roseum is distinctive from other species of the genus Trichothecium in its characteristic zigzag patterned chained conidia. It is found in various countries worldwide and can grow in a variety of habitats ranging from leaf litter to fruit crops. Trichothecium roseum produces a wide variety of secondary metabolites including mycotoxins, such as roseotoxins and trichothecenes, which can infect and spoil a variety of fruit crops. It can act as both a secondary and opportunistic pathogen by causing pink rot on various fruits and vegetables and thus has an economical impact on the farming industry. Secondary metabolites of T. roseum, specifically Trichothecinol A, are being investigated as potential anti-metastatic drugs. Several agents including harpin, silicon oxide, and sodium silicate are potential inhibitors of T. roseum growth on fruit crops. Trichothecium roseum is mainly a plant pathogen and has yet to show a significant impact on human health.

Harposporium anguillulae is a member of the genus Harposporium. It is an endoparasitic nematophagous fungus that attacks nematodes and eelworms and is isolated commonly from field and agricultural soils as well as used as an experimental organism in the laboratory.

Aspergillus clavatus is a species of fungus in the genus Aspergillus with conidia dimensions 3–4.5 x 2.5–4.5 μm. It is found in soil and animal manure. The fungus was first described scientifically in 1834 by the French mycologist John Baptiste Henri Joseph Desmazières.
Dactylellina haptotyla is a common soil-living fungus that develops structures to capture nematodes as nutrient source. In the presence of nematodes, spores can germinate into sticky knobs or non-constricting loops. The fungus traps nematodes with sticky knobs and non-constricting loops, then breakdown the cuticle, and penetrates the body of nematodes to obtain nutrients. For its predatory nature, Dactylellina haptotyla is also considered as nematode-trapping fungus or carnivorous fungus.
Pithomyces chartarum is a fungus predominantly found in subtropical countries and other localities with warmer climates. However, it occurs throughout the world including the United Kingdom, Europe and Netherlands. Pithomyces chartarum produces a mycotoxin called sporidesmin when it grows on plants, particularly grasses. Presence of the toxin in forage grasses causes facial eczema in sheep, and is especially problematic in areas such as New Zealand where sheep are intensively raised. Other health effect of P. chartarum are not well understood.
Arthrobotrys dactyloides is a species of fungus in the family Orbiliaceae. It is nematophagous, forming loops of hypha to trap nematodes, on which it then feeds.
Stylopage is a polytypic genus of predacious fungus in the order Zoopagales, within the subphylum Zoopagomycotina. All known species of Stylopage subsist on various species of amoebae or nematodes by trapping their prey, typically using an adhesive substance that coats their vegetative hyphae, and absorbing nutrients through the projection of a haustorium. 17 extant Stylopage species have been described thus far.
Birgit Ann-Marie Margareta Nordbring-Hertz, was a Swedish scientist at Lund University known for her work on the interactions between fungi and nematodes.

This glossary of mycology is a list of definitions of terms and concepts relevant to mycology, the study of fungi. Terms in common with other fields, if repeated here, generally focus on their mycology-specific meaning. Related terms can be found in glossary of biology and glossary of botany, among others. List of Latin and Greek words commonly used in systematic names and Botanical Latin may also be relevant, although some prefixes and suffixes very common in mycology are repeated here for clarity.
Meristacrum is a fungal genus in the monotypic family Meristacraceae, of the order Entomophthorales. They are parasites of soil invertebrates, they typically infect nematodes, and tardigrades.
Arthrobotrys musiformis is a species of nematode catching fungi, genus Arthrobotrys. This, like other Arthrobotrys species, captures and feeds on nematodes. It is widespread, with its initial discovery being in Norfolk, Virginia. This species demonstrates promising anti-helminth potential, and is hypothesized to reduce the number of parasitic nematodes in plants and livestock as either a biocontrol or through isolating metabolites.
