
An extinction event is a widespread and rapid decrease in the biodiversity on Earth. Such an event is identified by a sharp fall in the diversity and abundance of multicellular organisms. It occurs when the rate of extinction increases with respect to the background extinction rate and the rate of speciation. Estimates of the number of major mass extinctions in the last 540 million years range from as few as five to more than twenty. These differences stem from disagreement as to what constitutes a "major" extinction event, and the data chosen to measure past diversity.
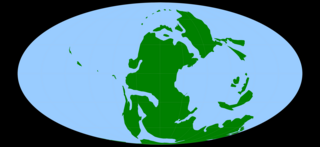
The Permian is a geologic period and stratigraphic system which spans 47 million years from the end of the Carboniferous Period 298.9 million years ago (Mya), to the beginning of the Triassic Period 251.902 Mya. It is the last period of the Paleozoic Era; the following Triassic Period belongs to the Mesozoic Era. The concept of the Permian was introduced in 1841 by geologist Sir Roderick Murchison, who named it after the region of Perm in Russia.
Approximately 251.9 million years ago, the Permian–Triassicextinction event forms the boundary between the Permian and Triassic geologic periods, and with them the Paleozoic and Mesozoic eras. It is the Earth's most severe known extinction event, with the extinction of 57% of biological families, 83% of genera, 81% of marine species and 70% of terrestrial vertebrate species. It is also the greatest known mass extinction of insects. It is the greatest of the "Big Five" mass extinctions of the Phanerozoic. There is evidence for one to three distinct pulses, or phases, of extinction.
The Late Ordovician mass extinction (LOME), sometimes known as the end-Ordovician mass extinction or the Ordovician-Silurian extinction, is the first of the "big five" major mass extinction events in Earth's history, occurring roughly 445 million years ago (Ma). It is often considered to be the second-largest known extinction event, in terms of the percentage of genera that became extinct. Extinction was global during this interval, eliminating 49–60% of marine genera and nearly 85% of marine species. Under most tabulations, only the Permian-Triassic mass extinction exceeds the Late Ordovician mass extinction in biodiversity loss. The extinction event abruptly affected all major taxonomic groups and caused the disappearance of one third of all brachiopod and bryozoan families, as well as numerous groups of conodonts, trilobites, echinoderms, corals, bivalves, and graptolites. Despite its taxonomic severity, the Late Ordovician mass extinction did not produce major changes to ecosystem structures compared to other mass extinctions, nor did it lead to any particular morphological innovations. Diversity gradually recovered to pre-extinction levels over the first 5 million years of the Silurian period.
The Late Devonian extinction consisted of several extinction events in the Late Devonian Epoch, which collectively represent one of the five largest mass extinction events in the history of life on Earth. The term primarily refers to a major extinction, the Kellwasser event, also known as the Frasnian-Famennian extinction, which occurred around 372 million years ago, at the boundary between the Frasnian stage and the Famennian stage, the last stage in the Devonian Period. Overall, 19% of all families and 50% of all genera became extinct. A second mass extinction called the Hangenberg event, also known as the end-Devonian extinction, occurred 359 million years ago, bringing an end to the Famennian and Devonian, as the world transitioned into the Carboniferous Period.
The Guadalupian is the second and middle series/epoch of the Permian. The Guadalupian was preceded by the Cisuralian and followed by the Lopingian. It is named after the Guadalupe Mountains of New Mexico and Texas, and dates between 272.95 ± 0.5 – 259.1 ± 0.4 Mya. The series saw the rise of the therapsids, a minor extinction event called Olson's Extinction and a significant mass extinction called the end-Capitanian extinction event. The Guadalupian was previously known as the Middle Permian.
The Lopingian is the uppermost series/last epoch of the Permian. It is the last epoch of the Paleozoic. The Lopingian was preceded by the Guadalupian and followed by the Early Triassic.
In the geologic timescale, the Capitanian is an age or stage of the Permian. It is also the uppermost or latest of three subdivisions of the Guadalupian Epoch or Series. The Capitanian lasted between 264.28 and 259.51 million years ago. It was preceded by the Wordian and followed by the Wuchiapingian.

The Early Triassic is the first of three epochs of the Triassic Period of the geologic timescale. It spans the time between 251.9 Ma and 247.2 Ma. Rocks from this epoch are collectively known as the Lower Triassic Series, which is a unit in chronostratigraphy. The Early Triassic is the oldest epoch of the Mesozoic Era. It is preceded by the Lopingian Epoch and followed by the Middle Triassic Epoch. The Early Triassic is divided into the Induan and Olenekian ages. The Induan is subdivided into the Griesbachian and Dienerian subages and the Olenekian is subdivided into the Smithian and Spathian subages.

In the geologic timescale, the Olenekian is an age in the Early Triassic epoch; in chronostratigraphy, it is a stage in the Lower Triassic series. It spans the time between 251.2 Ma and 247.2 Ma. The Olenekian is sometimes divided into the Smithian and the Spathian subages or substages. The Olenekian follows the Induan and is followed by the Anisian.

The Beaufort Group is the third of the main subdivisions of the Karoo Supergroup in South Africa. It is composed of a lower Adelaide Subgroup and an upper Tarkastad Subgroup. It follows conformably after the Ecca Group and unconformably underlies the Stormberg Group. Based on stratigraphic position, lithostratigraphic and biostratigraphic correlations, palynological analyses, and other means of geological dating, the Beaufort Group rocks are considered to range between Middle Permian (Wordian) to Early Triassic (Anisian) in age.

Anteosaurus is an extinct genus of large carnivorous dinocephalian synapsid. It lived at the end of the Guadalupian during the Capitanian stage, about 265 to 260 million years ago in what is now South Africa. It is mainly known by cranial remains and few postcranial bones. Measuring 5–6 m (16–20 ft) long and weighing about 600 kg (1,300 lb), Anteosaurus was the largest known carnivorous non-mammalian synapsid and the largest terrestrial predator of the Permian period. Occupying the top of the food chain in the Middle Permian, its skull, jaws and teeth show adaptations to capture large prey like the giants titanosuchids and tapinocephalids dinocephalians and large pareiasaurs.
The Daptocephalus Assemblage Zone is a tetrapod assemblage zone or biozone found in the Adelaide Subgroup of the Beaufort Group, a majorly fossiliferous and geologically important geological Group of the Karoo Supergroup in South Africa. This biozone has outcrops located in the upper Teekloof Formation west of 24°E, the majority of the Balfour Formation east of 24°E, and the Normandien Formation in the north. It has numerous localities which are spread out from Colesberg in the Northern Cape, Graaff-Reniet to Mthatha in the Eastern Cape, and from Bloemfontein to Harrismith in the Free State. The Daptocephalus Assemblage Zone is one of eight biozones found in the Beaufort Group and is considered Late Permian (Lopingian) in age. Its contact with the overlying Lystrosaurus Assemblage Zone marks the Permian-Triassic boundary.
The Emeishan Traps constitute a flood basalt volcanic province, or large igneous province, in south-western China, centred in Sichuan province. It is sometimes referred to as the Permian Emeishan Large Igneous Province or Emeishan Flood Basalts. Like other volcanic provinces or "traps", the Emeishan Traps are multiple layers of igneous rock laid down by large mantle plume volcanic eruptions. The Emeishan Traps eruptions were serious enough to have global ecological and paleontological impact.

The Abrahamskraal Formation is a geological formation and is found in numerous localities in the Northern Cape, Western Cape, and the Eastern Cape of South Africa. It is the lowermost formation of the Adelaide Subgroup of the Beaufort Group, a major geological group that forms part of the greater Karoo Supergroup. It represents the first fully terrestrial geological deposits of the Karoo Basin. Outcrops of the Abrahamskraal Formation are found from the small town Middelpos in its westernmost localities, then around Sutherland, the Moordenaarskaroo north of Laingsburg, Williston, Fraserburg, Leeu-Gamka, Loxton, and Victoria West in the Western Cape and Northern Cape. In the Eastern Cape outcrops are known from Rietbron, north of Klipplaat and Grahamstown, and also southwest of East London.

Pampaphoneus is an extinct genus of carnivorous dinocephalian therapsid belonging to the family Anteosauridae. It lived 268 to 265 million years ago during the Wordian age of the Guadalupian period in what is now Brazil. Pampaphoneus is known by an almost complete skull with the lower jaw still articulated, discovered on the lands of the Boqueirão Farm, near the city of São Gabriel, in the state of Rio Grande do Sul. A second specimen from the same locality was reported in 2019 and 2020 but has not yet been described. It is composed of a skull associated with postcranial remains. It is the first South American species of dinocephalian to have been described. The group was previously known in South America only by a few isolated teeth and a jaw fragment reported in 2000 in the same region of Brazil. Phylogenetic analysis conducted by Cisneros and colleagues reveals that Pampaphoneus is closely related to anteosaurs from European Russia, indicating a closer faunal relationship between South America and Eastern Europe than previously thought, thus promoting a Pangea B continental reconstruction.

The Middleton Formation is a geological formation that extends through the Northern Cape, Western Cape, and Eastern Cape provinces of South Africa. It overlies the lower Abrahamskraal Formation, and is the eastern correlate, East of 24ºE, of the Teekloof Formation. Outcrops and exposures of the Middleton Formation range from Graaff-Reinet in the Eastern Cape onwards. The Middleton Formation's type locality lies near the small hamlet, Middleton, approximately 25 km south of Cookhouse. Other exposures lie in hillsides along the Great Fish River in the Eastern Cape. The Middleton Formation forms part of the Adelaide Subgroup of the Beaufort Group, which itself forms part of the Karoo Supergroup.

Alatoconchidae is an extinct family of prehistoric bivalves that lived in the early to middle Permian period. Genera belonging to Alatoconchidae are characterized by their shell that is strongly compressed in the dorsoventral direction. Some species reached large sizes of as much as 1 metre (3.3 ft) long. It is hypothesized that some species in this family got energy from chemosynthetic bacteria.
The Toarcian extinction event, also called the Pliensbachian-Toarcian extinction event, the Early Toarcian mass extinction, the Early Toarcian palaeoenvironmental crisis, or the Jenkyns Event, was an extinction event that occurred during the early part of the Toarcian age, approximately 183 million years ago, during the Early Jurassic. The extinction event had two main pulses, the first being the Pliensbachian-Toarcian boundary event (PTo-E). The second, larger pulse, the Toarcian Oceanic Anoxic Event (TOAE), was a global oceanic anoxic event, representing possibly the most extreme case of widespread ocean deoxygenation in the entire Phanerozoic eon. In addition to the PTo-E and TOAE, there were multiple other, smaller extinction pulses within this span of time.
