
A chordate is an animal belonging to the phylum Chordata. All chordates possess, at some point during their larval or adult stages, five distinctive physical characteristics (synapomorphies) that distinguish them from other taxa. These five synapomorphies are a notochord, a hollow dorsal nerve cord, an endostyle or thyroid, pharyngeal slits, and a post-anal tail. The name "chordate" comes from the first of these synapomorphies, the notochord, which plays a significant role in chordate structure and movement. Chordates are also bilaterally symmetric, have a coelom, possess a circulatory system, and exhibit metameric segmentation.

Vetulicolia is a phylum of animals encompassing several extinct species belonging to the Cambrian Period. The phylum was created by Degan Shu and his research team in 2001, and named after Vetulicola cuneata, the first species of the phylum described in 1987. The vetulicolian body comprises two parts: a voluminous anterior forebody, tipped with an anteriorly positioned mouth and lined with a row of five round to oval-shaped features on each lateral side, which have been interpreted as gills ; and a posterior section that primitively comprises seven segments and functions as a tail. All vetulicolians lack preserved appendages of any kind, having no legs, feelers or even eyes. The area where the anterior and posterior parts join is constricted.

Crinoids are marine animals that make up the class Crinoidea. Crinoids that are attached to the sea bottom by a stalk in their juvenile form are commonly called sea lilies, while the unstalked forms, called feather stars or comatulids, are members of the largest crinoid order, Comatulida. Crinoids are echinoderms in the phylum Echinodermata, which also includes the starfish, brittle stars, sea urchins and sea cucumbers. They live in both shallow water and in depths as great as 9,000 meters (30,000 ft).

Helicoplacus is the earliest well-studied fossil echinoderm. Fossil plates are known from several regions. Complete specimens were found in Lower Cambrian strata of the White Mountains of California.

Cothurnocystis is a genus of small enigmatic echinoderms that lived during the Ordovician. Individual animals had a flat boot-shaped body and a thin rod-shaped appendage that may be a stem, or analogous to a foot or a tail. Fossils of Cothurnocystis species have been found in Nevada, Scotland, Czech Republic, France and Morocco.

Edrioasteroidea is an extinct class of echinoderms. The living animal would have resembled a pentamerously symmetrical disc or cushion. They were obligate encrusters and attached themselves to inorganic or biologic hard substrates. A 507 million years old species, Totiglobus spencensis, is actually the first known echinoderm adapted to live on a hard surface after the soft microbial mats that covered the seafloor were destroyed in the Cambrian substrate revolution.
The stylophorans are an extinct, possibly polyphyletic group allied to the Paleozoic Era echinoderms, comprising the prehistoric cornutes and mitrates. It is synonymous with the subphylum Calcichordata. Their unusual appearances have led to a variety of very different reconstructions of their anatomy, how they lived, and their relationships to other organisms.

Mitrates are an extinct group of stem group echinoderms, which may be closely related to the hemichordates. Along with the cornutes, they form one half of the Stylophora.

Deuterostomia are animals typically characterized by their anus forming before their mouth during embryonic development. The group's sister clade is Protostomia, animals whose digestive tract development is more varied. Some examples of deuterostomes include vertebrates, sea stars, and crinoids.

Camptostroma roddyi is an extinct echinoderm from the Bonnia-Olenellus Zone the Early Cambrian Kinzers Formation near York and Lancaster, Southeastern Pennsylvania. In life, it would have resembled a cupcake, with the axial skeleton forming a star pattern on the upper surface. It was originally thought, on the basis of its medusoid shape, to be a jellyfish-like organism, but the fossils themselves clearly rule out the possibility of a gelatinous body - the stereom plates are clearly preserved and possess the calcitic cleavage pattern diagnostic of echinoderms. It has been placed in a class of basal echinoderms, the Edrioasteroids.

Homalozoa is an obsolete extinct subphylum of Paleozoic era echinoderms, prehistoric marine invertebrates. They are also referred to as carpoids.

Stereom is a calcium carbonate material that makes up the internal skeletons found in all echinoderms, both living and fossilized forms. It is a sponge-like porous structure which, in a sea urchin may be 50% by volume living cells, and the rest being a matrix of calcite crystals. The size of openings in stereom varies in different species and in different places within the same organism. When an echinoderm becomes a fossil, microscopic examination is used to reveal the structure and such examination is often an important tool to classify the fossil as an echinoderm or related creature.
Diploporita is an extinct class of blastozoan that ranged from the Ordovician to the Devonian. These echinoderms are identified by a specialized respiratory structure, called diplopores. Diplopores are a double pore system that sit within a depression on a single thecal (body) plate; each plate can contain numerous diplopore pairs.
Paleontology or palaeontology is the study of prehistoric life forms on Earth through the examination of plant and animal fossils. This includes the study of body fossils, tracks (ichnites), burrows, cast-off parts, fossilised feces (coprolites), palynomorphs and chemical residues. Because humans have encountered fossils for millennia, paleontology has a long history both before and after becoming formalized as a science. This article records significant discoveries and events related to paleontology that occurred or were published in the year 2018.

Soluta is an extinct class of echinoderms that lived from the Middle Cambrian to the Early Devonian. The class is also known by its junior synonym Homoiostelea. Soluta is one of the four "carpoid" classes, alongside Ctenocystoidea, Cincta, and Stylophora, which made up the obsolete subphylum Homalozoa. Solutes were asymmetric animals with a stereom skeleton and two appendages, an arm extending anteriorly and a posterior appendage called a homoiostele.

Ctenocystoidea is an extinct clade of echinoderms, which lived during the Cambrian and Ordovician periods. Unlike other echinoderms, ctenocystoids had bilateral symmetry, or were only very slightly asymmetrical. They are believed to be one of the earliest-diverging branches of echinoderms, with their bilateral symmetry a trait shared with other deuterostomes. Ctenocystoids were once classified in the taxon Homalozoa, also known as Carpoidea, alongside cinctans, solutes, and stylophorans. Homalozoa is now recognized as a polyphyletic group of echinoderms without radial symmetry. Ctenocystoids were geographically widespread during the Middle Cambrian, with one species surviving into the Late Ordovician.
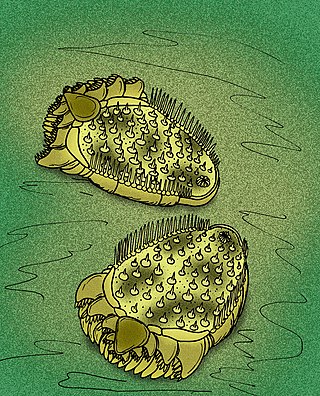
Ctenoimbricata is an extinct genus of bilaterally symmetrical echinoderm, which lived during the early Middle Cambrian period of what is now Spain. It contains one species, Ctenoimbricata spinosa. It may be the most basal known echinoderm. It resembles the extinct ctenocystoids and cinctans, particularly the basal ctenocystoid Courtessolea. Ctenoimbricata is interpreted as a deposit-feeding pharyngeal basket feeder. It was relatively small, with a body 20 millimetres (0.79 in) long.

Yanjiahella biscarpa is an extinct species of Ediacaran and Early Cambrian deuterostome which may represent the earliest stem group echinoderms.
Yorkicystis is a genus of edrioasteroid echinoderm that lived 510 million years ago in the Cambrian aged Kinzers Formation in what is now Pennsylvania. This genus is important as it provides some of the oldest evidence of echinoderms losing their hard mineralized outer skeletons. Yorkicystis also shows that some echinoderms lost their skeletons during the Cambrian, which is a greatly different time as to when most other species lost theirs.

Lepidocystis is a Palaeozoic genus of imbricate lepidocystoid eocrinoid, closely related to Kinzercystis, It bore a stalk, with which it attached to firm substrates; and brachioles arising as lateral branches from its arms.
