
Cnidaria is a phylum under kingdom Animalia containing over 11,000 species of aquatic animals found both in fresh water and marine environments, including jellyfish, hydroids, sea anemones, corals and some of the smallest marine parasites. Their distinguishing features are a decentralized nervous system distributed throughout a gelatinous body and the presence of cnidocytes or cnidoblasts, specialized cells with ejectable flagella used mainly for envenomation and capturing prey. Their bodies consist of mesoglea, a non-living, jelly-like substance, sandwiched between two layers of epithelium that are mostly one cell thick. Cnidarians are also some of the only animals that can reproduce both sexually and asexually.
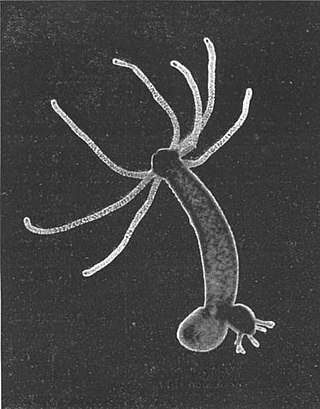
Hydra is a genus of small freshwater hydrozoans of the phylum Cnidaria. They are native to the temperate and tropical regions. The genus was named by Linnaeus in 1758 after the Hydra, which was the many-headed beast of myth defeated by Heracles, as when the animal has a part severed, it will regenerate much like the mythical hydra’s heads. Biologists are especially interested in Hydra because of their regenerative ability; they do not appear to die of old age, or to age at all.

Venom or zootoxin is a type of toxin produced by an animal that is actively delivered through a wound by means of a bite, sting, or similar action. The toxin is delivered through a specially evolved venom apparatus, such as fangs or a stinger, in a process called envenomation. Venom is often distinguished from poison, which is a toxin that is passively delivered by being ingested, inhaled, or absorbed through the skin, and toxungen, which is actively transferred to the external surface of another animal via a physical delivery mechanism.

Jellyfish, also known as sea jellies, are the medusa-phase of certain gelatinous members of the subphylum Medusozoa, which is a major part of the phylum Cnidaria.

Box jellyfish are cnidarian invertebrates distinguished by their box-like body. Some species of box jellyfish produce potent venom delivered by contact with their tentacles. Stings from some species, including Chironex fleckeri, Carukia barnesi, Malo kingi, and a few others, are extremely painful and often fatal to humans.

In zoology, a tentacle is a flexible, mobile, and elongated organ present in some species of animals, most of them invertebrates. In animal anatomy, tentacles usually occur in one or more pairs. Anatomically, the tentacles of animals work mainly like muscular hydrostats. Most forms of tentacles are used for grasping and feeding. Many are sensory organs, variously receptive to touch, vision, or to the smell or taste of particular foods or threats. Examples of such tentacles are the eyestalks of various kinds of snails. Some kinds of tentacles have both sensory and manipulatory functions.
Chironex fleckeri, commonly known as the Australian box jelly, and nicknamed the sea wasp, is a species of extremely venomous box jellyfish found in coastal waters from northern Australia and New Guinea to Indonesia, Cambodia, Malaysia and Singapore, the Philippines and Vietnam. It has been described as "the most lethal jellyfish in the world", with at least 64 known deaths in Australia from 1884 to 2021.

Medusozoa is a clade in the phylum Cnidaria, and is often considered a subphylum. It includes the classes Hydrozoa, Scyphozoa, Staurozoa and Cubozoa, and possibly the parasitic Polypodiozoa. Medusozoans are distinguished by having a medusa stage in their often complex life cycle, a medusa typically being an umbrella-shaped body with stinging tentacles around the edge. With the exception of some Hydrozoa, all are called jellyfish in their free-swimming medusa phase.

Craspedacusta sowerbii or peach blossom jellyfish is a species of freshwater hydrozoan jellyfish, or hydromedusa cnidarian. Hydromedusan jellyfish differ from scyphozoan jellyfish because they have a muscular, shelf-like structure called a velum on the ventral surface, attached to the bell margin. Originally from the Yangtze basin in China, C. sowerbii is an introduced species now found throughout the world in bodies of fresh water.

The starlet sea anemone is a species of small sea anemone in the family Edwardsiidae native to the east coast of the United States, with introduced populations along the coast of southeast England and the west coast of the United States. Populations have also been located in Nova Scotia, Canada. This sea anemone is found in the shallow brackish water of coastal lagoons and salt marshes where its slender column is usually buried in the mud and its tentacles exposed. Its genome has been sequenced and it is cultivated in the laboratory as a model organism, but the IUCN has listed it as being a "Vulnerable species" in the wild.

Pelagia noctiluca is a jellyfish in the family Pelagiidae and the only currently recognized species in the genus Pelagia. It is typically known in English as the mauve stinger, but other common names are purple-striped jelly, purple stinger, purple people eater, purple jellyfish, luminous jellyfish and night-light jellyfish. In Greek, pelagia means "(she) of the sea", from pelagos "sea, open sea"; in Latin noctiluca is the combining form of nox, "night"", and lux, "light"; thus, Pelagia noctiluca can be described as a marine organism with the ability to glow in the dark (bioluminescence). It is found worldwide in tropical and warm temperate seas, although it is suspected that records outside the North Atlantic region, which includes the Mediterranean and Gulf of Mexico, represent closely related but currently unrecognized species.

Sea anemones are a group of predatory marine invertebrates constituting the order Actiniaria. Because of their colourful appearance, they are named after the Anemone, a terrestrial flowering plant. Sea anemones are classified in the phylum Cnidaria, class Anthozoa, subclass Hexacorallia. As cnidarians, sea anemones are related to corals, jellyfish, tube-dwelling anemones, and Hydra. Unlike jellyfish, sea anemones do not have a medusa stage in their life cycle.

Condylactis gigantea is a tropical species of ball anemone that is found in shallow reefs and other shallow inshore areas in the Caribbean Sea – more specifically the West Indies – and the western Atlantic Ocean including southern Florida through the Florida Keys. It is also commonly known as: giant Caribbean sea anemone, giant golden anemone, condylactis anemone, Haitian anemone, pink-tipped anemone, purple-tipped anemone, and Florida condy. This species can easily be seen growing in lagoons or in inner reefs as either individuals or loose groups, but never as colonies. They are often used as a model organism along with others in their genus for facultative symbiosis with monocellular algae.

A cnidosac is an anatomical feature that is found in the group of sea slugs known as aeolid nudibranchs, a clade of marine opisthobranch gastropod molluscs. A cnidosac contains cnidocytes, stinging cells that are also known as cnidoblasts or nematocysts. These stinging cells are not made by the nudibranch, but by the species that it feeds upon. However, once the nudibranch is armed with these stinging cells, they are used in its own defense.

The Enthemonae is a suborder of sea anemones in the order Actiniaria. It comprises those sea anemones with typical arrangement of mesenteries for actiniarians.

A nematocyst is a subcellular structure or organelle containing extrusive filaments found in two families of athecate dinoflagellates, the Warnowiaceae and Polykrikaceae. It is distinct from the similar subcellular structures found in the cnidocyte cells of cnidarians, a group of multicellular organisms including jellyfish and corals; such structures are also often called nematocysts, and cnidocytes are sometimes referred to as nematocytes. It is unclear whether the relationship between dinoflagellate and cnidarian nematocysts is a case of convergent evolution or common descent, although molecular evidence has been interpreted as supporting an endosymbiotic origin for cnidarian nematocysts.
Nematosomes are multicellular motile bodies found in the gastrovascular cavity of the model sea anemone Nematostella vectensis starlet sea anemone. First described by Stephenson in 1935, nematosomes are the defining apomorphy (synapomorphy) of the genus Nematostella but have received relatively little study. Nematosomes can be observed circulating through the body cavity and tentacle lumen of adult anemones, occasionally coming to rest on the gastrodermis. Nematosomes that are dislodged from rest return to circulation. The lifespan of a single nematosome has not been studied.

The thimble jellyfish is a species of cnidarian found in the warm West Atlantic Ocean, including the Caribbean. It is a tiny jellyfish with a straight-sided, flat-topped bell. This jellyfish is the most common cause of seabather's eruption, a reaction caused by the injection of juvenile jellyfish nematocysts into human skin.

Aiptasia mutabilis, also known as the Trumpet anemone, Rock anemone, and Glass anemone, is a species of anemone typically found attached to substrates in cold waters of the Atlantic Ocean. Its unique trumpet shape gives it its common name and it can grow to be 12 cm, having a column between 3 and 6 cm in size. Like many cnidarians, they rely on nematocysts for protection and to capture prey. They are not difficult to care for, and can be kept in a home aquarium, although due to their speed of reproduction, can quickly become overpopulated.

Clytia hemisphaerica is a small hydrozoan-group cnidarian, about 1 cm in diameter, that is found in the Mediterranean Sea and the North-East Atlantic Ocean. Clytia has the free-swimming jellyfish form typical of the Hydrozoa, as well as vegetatively propagating polyps.




