Countergradient is a type of phenotypic plasticity
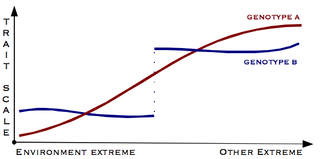
Countergradient variation is a type of phenotypic plasticity that occurs when the phenotypic variation determined by a biological population's genetic components opposes the phenotypic variation caused by an environmental gradient.[1][2] This can cause different populations of the same organism to display similar phenotypes regardless of their underlying genetics and differences in their environments.[1]
To illustrate a common example known as countergradient growth rate, consider two populations. The two populations live in different environments that affect growth differently due to many ecological factors, such as the temperature and available food. One population is genetically predisposed to have an increased growth rate but inhabits an environment that reduces growth rate, such as a cool environment, and thereby limits the opportunities to take full advantage of any genetic predisposition. The second population is genetically predisposed to have a decreased growth rate but inhabits an environment that supports an increased growth rate, such as a warm environment, and allows members of the population to grow faster despite their genetic disadvantage. Since the genetic influence directly counteracts the environmental influence in each population, both populations will have a similar intermediate growth rate. Countergradient variation can reduce apparent variability by creating similar phenotypes, but it is still possible for the two populations to show phenotypic diversity if either the genetic gradient or the environmental gradient has a stronger influence.
Many examples of countergradient variation have been discovered through the use of transplant experiments. Countergradient variation of growth rate is one of the most common examples. Growth rate and body size have important ecological implications, such as how they impact an organism's survival, life history, and fecundity.[3] Countergradient variation has been described in many ectothermic animals, since ectotherms rely on environmental temperature to regulate their metabolic rates, and thus, their growth rates. Ectotherms grow at a slower rate as latitude increases due to this impact of temperature. However, under countergradient variation, when these same animals are placed in a common environment with their low-latitude relatives, they grow faster relative to the low-latitude population. These studies are useful in predicting how animals can adapt to and will survive in different environmental conditions.
History
Green frog Lithobates clamitans unexpectedly grows faster in montane populations
Countergradient variation, originally termed "contra-gradient variation", was coined by Richard Levins in his 1968 book Evolution in Changing Environments.[2] Levins first used the term when describing patterns of body size across an altitudinal gradient in populations of Drosophila,[4] and since then many other instances of countergradient variation have been discovered. A study by Keith Berven and others was the first to find countergradient variation in a vertebrate.[5] Specifically, they found that in comparing montane, high altitude, populations to lowland, low altitude populations of the green frog (Lithobates clamitans, formerly Rana clamitans), rates of growth and development were higher in the montane populations. This is counter to what is expected since high altitude populations grow slower than low altitude ones in their respective environments. Similar to the results of this study, most of the known instances of countergradient variation are associated with a latitudinal or altitudinal gradient having an effect on growth rate (see Examples section).
Examples
The following are among the discovered cases of countergradient variation. The species name is followed by the trait affected and the environmental gradient studied.
Countergradient variation is the opposite of cogradient variation, in which the environmental effect on the phenotype enhances the genetic effect on the phenotype.[2] One common way to test for both of these patterns is with transplant experiments.[1] By bringing the two populations to the same environment, the environmental effect on phenotype is eliminated and only the genetic effect will cause variation. If cogradient variation is occurring, the same relationship will be seen in the common environment as the two natural environments. However, if countergradient variation is occurring the opposite relationship of what is seen in the natural environments will be seen in the common environment.[13] For example, if Population 1 has higher growth than Population 2 in their respective natural environments, countergradient variation can be detected if, when brought to the same environment, Population 1 now has lower growth than Population 2. Many of the examples listed above were discovered through these types of experiments. However, the mechanisms of how these differential growth rates arise is not fully understood.
Take the example of Sceloporus undulatus, the eastern fence lizard. These lizards demonstrate countergradient variation in growth rate across a latitudinal gradient. That is, in a common environment eggs from populations from the northern part of their range hatch out sooner than the eggs from populations at a more southern range.[8] This is opposite the pattern seen in their native habitats. One explanation for this could be that the northern eggs are also naturally larger, so they have more yolk energy available. Originally this maternal investment in larger eggs was thought to support increased developmental rate. However when yolk is removed from the eggs so that the populations have equal energy sources, there is still a substantial difference in growth.[8] Therefore, the mechanism behind this differential growth rate may lie in the efficiency of energy use.[14]
Ecological relevance
Populations that inhabit high latitudes experience shorter growing seasons than those that inhabit low latitudes due to the differences in seasonality. Because of this, it is thought that countergradient variation of growth is a means to compensate for the short amount of time juveniles have to prepare for winter.[13] With a genetic disposition to grow faster, these individuals can reach a large enough body size to survive through the winter. Increased body size is also associated with higher reproductive output.[3] Additionally, when countergradient variation acts on developmental rates, embryos that develop sooner or hatch out larger will have more time to grow or require less resources, respectively.
Since these inherently fast growth rates are not observed in all populations, it seems there are potential trade-offs that would prevent this from being beneficial in low latitude populations. One proposed detriment of enhanced embryonic growth is that animals use too much of their supplied nutrients or yolk during development. In some species of lizards the yolk remaining after hatching is absorbed into the body and used as energy for growth as a hatchling.[15] The use of more yolk energy during development diminishes the reserves available for hatchling growth and can impact hatchling survival.[15][16] Another hypothesis is that animals that grow quickly do not expend as much energy on differentiation or cellular maintenance because they have shorter incubation times.[8] This allows more energy to be used for growth, but less energy for differentiation. Increased growth has been shown to lead to higher occurrences of defects or complications that cause higher rates of mortality. Some examples include rapid growth leading to bone deformities in sandhill cranes[17] and increased risk of cardiovascular problems in Atlantic salmon.[18]
Animals are able to employ different strategies for growth and development to counteract environmental challenges. However, the fact that all individuals do not grow faster and the presence of the mentioned potential fitness trade-offs show that there must be some limit to how much animals can use countergradient growth to compensate for environmental conditions that cause slow growth. Studies of countergradient variation are being explored as a useful way to predict the evolutionary constraints animals face in differing environmental conditions.
Notes
1 2 3 Conover, David; Schultz, Eric (1995). "Phenotypic similarity and the evolutionary significance of countergradient variation". Trends in Ecology & Evolution. 10 (6): 248–252. doi:10.1016/s0169-5347(00)89081-3. PMID21237029.
1 2 3 Levins, Richard (1968). Evolution in Changing Environments. Princeton University Press.
1 2 Yampolsky, Lev Y.; Scheiner, Samuel M. (1996). "Why Larger Offspring at Lower Temperatures? A Demographic Approach". The American Naturalist. 147 (1): 86–100. doi:10.1086/285841. ISSN0003-0147. S2CID84821433.
1 2 Levins, Richard = (1969). "Thermal Acclimation and Heat Resistance in Drosophila Species". The American Naturalist. 103 (933): 483–499. doi:10.1086/282616. JSTOR2459410. S2CID84404474.
1 2 Berven, Keith A.; Gill, Douglas E.; Smith-Gill, Sandra J. (1979). "Countergradient Selection in the Green Frog, Rana clamitans". Evolution. 33 (2): 609–623. doi:10.2307/2407784. JSTOR2407784. PMID28563934.
↑ Berven, Keith A. (1982). "The Genetic Basis of Altitudinal Variation in the Wood Frog Rana sylvatica. I. An Experimental Analysis of Life History Traits". Evolution. 36 (5): 962–983. doi:10.2307/2408075. JSTOR2408075. PMID28567824.
↑ Gerard, V. A.; Bois, K. R. Du (1988). "Temperature ecotypes near the southern boundary of the kelp Laminaria saccharina". Marine Biology. 97 (4): 575–580. doi:10.1007/BF00391054. S2CID83539774.
↑ Chapin, F. Stuart; Chapin, Melissa C. (1981). "Ecotypic Differentiation of Growth Processes in Carex Aquatilis along Latitudinal and Local Gradients". Ecology. 62 (4): 1000–1009. doi:10.2307/1936999. ISSN1939-9170. JSTOR1936999.
1 2 Troyer, Katherine (1987). "Posthatching Yolk in a Lizard: Internalization and Contribution to Growth". Journal of Herpetology. 21 (2): 102–106. doi:10.2307/1564470. JSTOR1564470.
↑ Sinervo, Barry (1990). "The Evolution of Maternal Investment in Lizards: An Experimental and Comparative Analysis of Egg Size and Its Effects on Offspring Performance". Evolution. 44 (2): 279–294. doi:10.2307/2409407. JSTOR2409407. PMID28564384.
↑ Serafin, J. A. (1982-01-01). "The Influence of Diet Composition upon Growth and Development of Sandhill Cranes". The Condor. 84 (4): 427–434. doi:10.2307/1367448. JSTOR1367448.
↑ Saunders, R. L.; Farrell, A. P.; Knox, D. E. (1992). "Progression of Coronary Arterial Lesions in Atlantic Salmon (Salmo salar) as a Function of Growth Rate". Canadian Journal of Fisheries and Aquatic Sciences. 49 (5): 878–884. doi:10.1139/f92-098.
GigantothermyLarge ectothermic animals more easily maintain constant body temperature
Related Research Articles
In genetics, the phenotype is the set of observable characteristics or traits of an organism. The term covers the organism's morphology or physical form and structure, its developmental processes, its biochemical and physiological properties, its behavior, and the products of behavior. An organism's phenotype results from two basic factors: the expression of an organism's genetic code, or its genotype, and the influence of environmental factors. Both factors may interact, further affecting phenotype. When two or more clearly different phenotypes exist in the same population of a species, the species is called polymorphic. A well-documented example of polymorphism is Labrador Retriever coloring; while the coat color depends on many genes, it is clearly seen in the environment as yellow, black, and brown. Richard Dawkins in 1978 and then again in his 1982 book The Extended Phenotype suggested that one can regard bird nests and other built structures such as caddis-fly larva cases and beaver dams as "extended phenotypes".
A maternal effect is a situation where the phenotype of an organism is determined not only by the environment it experiences and its genotype, but also by the environment and genotype of its mother. In genetics, maternal effects occur when an organism shows the phenotype expected from the genotype of the mother, irrespective of its own genotype, often due to the mother supplying messenger RNA or proteins to the egg. Maternal effects can also be caused by the maternal environment independent of genotype, sometimes controlling the size, sex, or behaviour of the offspring. These adaptive maternal effects lead to phenotypes of offspring that increase their fitness. Further, it introduces the concept of phenotypic plasticity, an important evolutionary concept. It has been proposed that maternal effects are important for the evolution of adaptive responses to environmental heterogeneity.
Bergmann's rule is an ecogeographical rule that states that within a broadly distributed taxonomic clade, populations and species of larger size are found in colder environments, while populations and species of smaller size are found in warmer regions. Bergmann's rule only describes the overall size of the animals, but does not include body parts like Allen's rule does.
An environmental gradient, or climate gradient, is a change in abiotic (non-living) factors through space. Environmental gradients can be related to factors such as altitude, depth, temperature, soil humidity and precipitation. Often times, a multitude of biotic (living) factors are closely related to these gradients; as a result of a change in an environmental gradient, factors such as species abundance, population density, morphology, primary productivity, predation, and local adaptation may be impacted.
Genetic architecture is the underlying genetic basis of a phenotypic trait and its variational properties. Phenotypic variation for quantitative traits is, at the most basic level, the result of the segregation of alleles at quantitative trait loci (QTL). Environmental factors and other external influences can also play a role in phenotypic variation. Genetic architecture is a broad term that can be described for any given individual based on information regarding gene and allele number, the distribution of allelic and mutational effects, and patterns of pleiotropy, dominance, and epistasis.
Evolutionary capacitance is the storage and release of variation, just as electric capacitors store and release charge. Living systems are robust to mutations. This means that living systems accumulate genetic variation without the variation having a phenotypic effect. But when the system is disturbed, robustness breaks down, and the variation has phenotypic effects and is subject to the full force of natural selection. An evolutionary capacitor is a molecular switch mechanism that can "toggle" genetic variation between hidden and revealed states. If some subset of newly revealed variation is adaptive, it becomes fixed by genetic assimilation. After that, the rest of variation, most of which is presumably deleterious, can be switched off, leaving the population with a newly evolved advantageous trait, but no long-term handicap. For evolutionary capacitance to increase evolvability in this way, the switching rate should not be faster than the timescale of genetic assimilation.
The common side-blotched lizard is a species of side-blotched lizard in the family Phrynosomatidae. The species is native to dry regions of the western United States and northern Mexico. It is notable for having a unique form of polymorphism wherein each of the three different male morphs utilizes a different strategy in acquiring mates. The three morphs compete against each other following a pattern of rock paper scissors, where one morph has advantages over another but is outcompeted by the third.
A polyphenic trait is a trait for which multiple, discrete phenotypes can arise from a single genotype as a result of differing environmental conditions. It is therefore a special case of phenotypic plasticity.
Phenotypic plasticity refers to some of the changes in an organism's behavior, morphology and physiology in response to a unique environment. Fundamental to the way in which organisms cope with environmental variation, phenotypic plasticity encompasses all types of environmentally induced changes that may or may not be permanent throughout an individual's lifespan.
Canalisation is a measure of the ability of a population to produce the same phenotype regardless of variability of its environment or genotype. It is a form of evolutionary robustness. The term was coined in 1942 by C. H. Waddington to capture the fact that "developmental reactions, as they occur in organisms submitted to natural selection...are adjusted so as to bring about one definite end-result regardless of minor variations in conditions during the course of the reaction". He used this word rather than robustness to take into account that biological systems are not robust in quite the same way as, for example, engineered systems.
Genetic assimilation is a process described by Conrad H. Waddington by which a phenotype originally produced in response to an environmental condition, such as exposure to a teratogen, later becomes genetically encoded via artificial selection or natural selection. Despite superficial appearances, this does not require the (Lamarckian) inheritance of acquired characters, although epigenetic inheritance could potentially influence the result. Waddington stated that genetic assimilation overcomes the barrier to selection imposed by what he called canalization of developmental pathways; he supposed that the organism's genetics evolved to ensure that development proceeded in a certain way regardless of normal environmental variations.
In biology, a cline is a measurable gradient in a single character of a species across its geographical range. First coined by Julian Huxley in 1938, the "character" of the cline referred to is usually genetic, or phenotypic. Clines can show smooth, continuous gradation in a character, or they may show more abrupt changes in the trait from one geographic region to the next.
Evolutionary physiology is the study of the biological evolution of physiological structures and processes; that is, the manner in which the functional characteristics of individuals in a population of organisms have responded to natural selection across multiple generations during the history of the population. It is a sub-discipline of both physiology and evolutionary biology. Practitioners in the field come from a variety of backgrounds, including physiology, evolutionary biology, ecology, and genetics.
The eastern three-lined skink, also known as the bold-striped cool-skink, is a species of skink, a lizard in the family Scincidae. The species is endemic to Australia. A. duperreyi has been extensively studied in the context of understand the evolution of learning, viviparity in lizards, and temperature- and genetic-sex determination. A. duperreyi is classified as a species of least concern by the IUCN.
Anelosimus studiosus is a subsocial tangle web spider or theridiid spider living in both North America and South America. In 2012, genetic analysis revealed a previously identified species, A. tungurahua, is in fact the same species as A. studiosus.
Scathophaga stercoraria, commonly known as the yellow dung fly or the golden dung fly, is one of the most familiar and abundant flies in many parts of the Northern Hemisphere. As its common name suggests, it is often found on the feces of large mammals, such as horses, cattle, sheep, deer, and wild boar, where it goes to breed. The distribution of S. stercoraria is likely influenced by human agriculture, especially in northern Europe and North America. The Scathophaga are integral in the animal kingdom due to their role in the natural decomposition of dung in fields. They are also very important in the scientific world due to their short life cycles and susceptibility to experimental manipulations; thus, they have contributed significant knowledge about animal behavior.
Ecophenotypic variation ("ecophenotype") refers to phenotypical variation as a function of life station. In wide-ranging species, the contributions of heredity and environment are not always certain, but their interplay can sometimes be determined by experiment.
In evolutionary biology, developmental bias refers to the production against or towards certain ontogenetic trajectories which ultimately influence the direction and outcome of evolutionary change by affecting the rates, magnitudes, directions and limits of trait evolution. Historically, the term was synonymous with developmental constraint, however, the latter has been more recently interpreted as referring solely to the negative role of development in evolution.
The temperature-size rule denotes the plastic response of organismal body size to environmental temperature variation. Organisms exhibiting a plastic response are capable of allowing their body size to fluctuate with environmental temperature. First coined by David Atkinson in 1996, it is considered to be a unique case of Bergmann's rule that has been observed in plants, animals, birds, and a wide variety of ectotherms. Although exceptions to the temperature-size rule exist, recognition of this widespread "rule" has amassed efforts to understand the physiological mechanisms underlying growth and body size variation in differing environmental temperatures.
Urban evolution refers to the heritable genetic changes of populations in response to urban development and anthropogenic activities in urban areas. Urban evolution can be caused by mutation, genetic drift, gene flow, or evolution by natural selection. Biologists have observed evolutionary change in numerous species compared to their rural counterparts on a relatively short timescale.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.