
Nitrification is the biological oxidation of ammonia to nitrate via the intermediary nitrite. Nitrification is an important step in the nitrogen cycle in soil. The process of complete nitrification may occur through separate organisms or entirely within one organism, as in comammox bacteria. The transformation of ammonia to nitrite is usually the rate limiting step of nitrification. Nitrification is an aerobic process performed by small groups of autotrophic bacteria and archaea.
A hyperthermophile is an organism that thrives in extremely hot environments—from 60 °C (140 °F) upwards. An optimal temperature for the existence of hyperthermophiles is often above 80 °C (176 °F). Hyperthermophiles are often within the domain Archaea, although some bacteria are also able to tolerate extreme temperatures. Some of these bacteria are able to live at temperatures greater than 100 °C, deep in the ocean where high pressures increase the boiling point of water. Many hyperthermophiles are also able to withstand other environmental extremes, such as high acidity or high radiation levels. Hyperthermophiles are a subset of extremophiles. Their existence may support the possibility of extraterrestrial life, showing that life can thrive in environmental extremes.

In biochemistry, an ether lipid refers to any lipid in which the lipid "tail" group is attached to the glycerol backbone via an ether bond at any position. In contrast, conventional glycerophospholipids and triglycerides are triesters. Structural types include:

Sulfolobus is a genus of microorganism in the family Sulfolobaceae. It belongs to the archaea domain.

TEX86 is an organic paleothermometer based upon the membrane lipids of mesophilic marine Nitrososphaerota (formerly "Thaumarchaeota", "Marine Group 1 Crenarchaeota").

A thermoacidophile is an extremophilic microorganism that is both thermophilic and acidophilic; i.e., it can grow under conditions of high temperature and low pH. The large majority of thermoacidophiles are archaea or bacteria, though occasional eukaryotic examples have been reported. Thermoacidophiles can be found in hot springs and solfataric environments, within deep sea vents, or in other environments of geothermal activity. They also occur in polluted environments, such as in acid mine drainage.
Phytane is the isoprenoid alkane formed when phytol, a chemical substituent of chlorophyll, loses its hydroxyl group. When phytol loses one carbon atom, it yields pristane. Other sources of phytane and pristane have also been proposed than phytol.
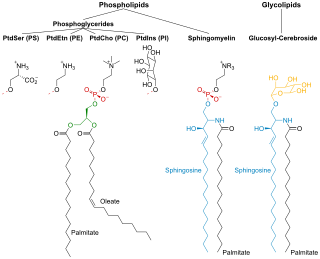
Membrane lipids are a group of compounds which form the lipid bilayer of the cell membrane. The three major classes of membrane lipids are phospholipids, glycolipids, and cholesterol. Lipids are amphiphilic: they have one end that is soluble in water ('polar') and an ending that is soluble in fat ('nonpolar'). By forming a double layer with the polar ends pointing outwards and the nonpolar ends pointing inwards membrane lipids can form a 'lipid bilayer' which keeps the watery interior of the cell separate from the watery exterior. The arrangements of lipids and various proteins, acting as receptors and channel pores in the membrane, control the entry and exit of other molecules and ions as part of the cell's metabolism. In order to perform physiological functions, membrane proteins are facilitated to rotate and diffuse laterally in two dimensional expanse of lipid bilayer by the presence of a shell of lipids closely attached to protein surface, called annular lipid shell.

Nitrosopumilus maritimus is an extremely common archaeon living in seawater. It is the first member of the Group 1a Nitrososphaerota to be isolated in pure culture. Gene sequences suggest that the Group 1a Nitrososphaerota are ubiquitous with the oligotrophic surface ocean and can be found in most non-coastal marine waters around the planet. It is one of the smallest living organisms at 0.2 micrometers in diameter. Cells in the species N. maritimus are shaped like peanuts and can be found both as individuals and in loose aggregates. They oxidize ammonia to nitrite and members of N. maritimus can oxidize ammonia at levels as low as 10 nanomolar, near the limit to sustain its life. Archaea in the species N. maritimus live in oxygen-depleted habitats. Oxygen needed for ammonia oxidation might be produced by novel pathway which generates oxygen and dinitrogen. N. maritimus is thus among organisms which are able to produce oxygen in dark.

Caldarchaeol is a membrane-spanning lipid of the glycerol dialkyl glycerol tetraether class. It is found in hyperthermophilic archaea. Membranes made up of caldarchaeol are more stable since the hydrophobic chains are linked together, allowing the microorganisms to withstand high temperatures. It is also known as dibiphytanyldiglycerol tetraether. Two glycerol units are linked together by two strains which consist of two phytanes linked together to form a linear chain of 32 carbon atoms.
Fuselloviridae is a family of viruses. Sulfolobus species, specifically shibatae, solfataricus, and islandicus, serve as natural hosts. There are two genera and nine species in the family. The Fuselloviridae are ubiquitous in high-temperature (≥70 °C), acidic hot springs around the world.

Archaea is a domain of single-celled organisms. These microorganisms lack cell nuclei and are therefore prokaryotes. Archaea were initially classified as bacteria, receiving the name archaebacteria, but this term has fallen out of use.
Archaeol is composed of two phytanyl chains linked to the sn-2 and sn-3 positions of glycerol. As its phosphate ester, it is a common component of the membranes of archaea.
The Nitrososphaerota are a phylum of the Archaea proposed in 2008 after the genome of Cenarchaeum symbiosum was sequenced and found to differ significantly from other members of the hyperthermophilic phylum Thermoproteota. Three described species in addition to C. symbiosum are Nitrosopumilus maritimus, Nitrososphaera viennensis, and Nitrososphaera gargensis. The phylum was proposed in 2008 based on phylogenetic data, such as the sequences of these organisms' ribosomal RNA genes, and the presence of a form of type I topoisomerase that was previously thought to be unique to the eukaryotes. This assignment was confirmed by further analysis published in 2010 that examined the genomes of the ammonia-oxidizing archaea Nitrosopumilus maritimus and Nitrososphaera gargensis, concluding that these species form a distinct lineage that includes Cenarchaeum symbiosum. The lipid crenarchaeol has been found only in Nitrososphaerota, making it a potential biomarker for the phylum. Most organisms of this lineage thus far identified are chemolithoautotrophic ammonia-oxidizers and may play important roles in biogeochemical cycles, such as the nitrogen cycle and the carbon cycle. Metagenomic sequencing indicates that they constitute ~1% of the sea surface metagenome across many sites.
Nitrososphaera is a mesophilic genus of ammonia-oxidizing Crenarchaeota. The first Nitrososphaera organism was discovered in garden soils at the University of Vienna leading to the categorization of a new genus, family, order and class of Archaea. This genus is contains three distinct species: N. viennensis, Ca. N. gargensis, and Ca N. evergladensis. Nitrososphaera are chemolithoautotrophs and have important biogeochemical roles as nitrifying organisms.
Nitrososphaera gargensis is a non-pathogenic, small coccus measuring 0.9 ± 0.3 μm in diameter. N. gargensis is observed in small abnormal cocci groupings and uses its archaella to move via chemotaxis. Being an Archaeon, Nitrososphaera gargensis has a cell membrane composed of crenarchaeol, its isomer, and a distinct glycerol dialkyl glycerol tetraether (GDGT), which is significant in identifying ammonia-oxidizing archaea (AOA). The organism plays a role in influencing ocean communities and food production.
Jessica E. Tierney (born 1982) is an American paleoclimatologist who has worked with geochemical proxies such as marine sediments, mud, and TEX86, to study past climate in East Africa. Her papers have been cited more than 2,500 times; her most cited work is Northern Hemisphere Controls on Tropical Southeast African Climate During the Past 60,000 Years. Tierney is currently an associate professor of geosciences and the Thomas R. Brown Distinguished Chair in Integrative Science at the University of Arizona and faculty affiliate in the University of Arizona School of Geography, Development and Environment Tierney is the first climatologist to win NSF's Alan T Waterman Award (2022) since its inception in 1975.
Paula Veronica Welander is a microbiologist and professor at Stanford University who is known for her research using lipid biomarkers to investigate how life evolved on Earth.
Glycerol dialkyl glycerol tetraether lipids (GDGTs) are a class of membrane lipids synthesized by archaea and some bacteria, making them useful biomarkers for these organisms in the geological record. Their presence, structure, and relative abundances in natural materials can be useful as proxies for temperature, terrestrial organic matter input, and soil pH for past periods in Earth history. Some structural forms of GDGT form the basis for the TEX86 paleothermometer. Isoprenoid GDGTs, now known to be synthesized by many archaeal classes, were first discovered in extremophilic archaea cultures. Branched GDGTs, likely synthesized by acidobacteriota, were first discovered in a natural Dutch peat sample in 2000.
Biphytane (or bisphytane) is a C40 isoprenoid produced from glycerol dialkyl glycerol tetraether (GDGT) degradation. As a common lipid membrane component, biphytane is widely used as a biomarker for archaea. In particular, given its association with sites of active anaerobic oxidation of methane (AOM), it is considered a biomarker of methanotrophic archaea. It has been found in both marine and terrestrial environments.

