
Toll-like receptors (TLRs) are a class of proteins that play a key role in the innate immune system. They are single-spanning receptors usually expressed on sentinel cells such as macrophages and dendritic cells, that recognize structurally conserved molecules derived from microbes. Once these microbes have reached physical barriers such as the skin or intestinal tract mucosa, they are recognized by TLRs, which activate immune cell responses. The TLRs include TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12, and TLR13. Humans lack genes for TLR11, TLR12 and TLR13 and mice lack a functional gene for TLR10. The receptors TLR1, TLR2, TLR4, TLR5, TLR6, and TLR10 are located on the cell membrane, whereas TLR3, TLR7, TLR8, and TLR9 are located in intracellular vesicles.

Defensins are small cysteine-rich cationic proteins across cellular life, including vertebrate and invertebrate animals, plants, and fungi. They are host defense peptides, with members displaying either direct antimicrobial activity, immune signaling activities, or both. They are variously active against bacteria, fungi and many enveloped and nonenveloped viruses. They are typically 18-45 amino acids in length, with three or four highly conserved disulphide bonds.

Antimicrobial peptides (AMPs), also called host defence peptides (HDPs) are part of the innate immune response found among all classes of life. Fundamental differences exist between prokaryotic and eukaryotic cells that may represent targets for antimicrobial peptides. These peptides are potent, broad spectrum antimicrobials which demonstrate potential as novel therapeutic agents. Antimicrobial peptides have been demonstrated to kill Gram negative and Gram positive bacteria, enveloped viruses, fungi and even transformed or cancerous cells. Unlike the majority of conventional antibiotics it appears that antimicrobial peptides frequently destabilize biological membranes, can form transmembrane channels, and may also have the ability to enhance immunity by functioning as immunomodulators.
The following are notable events in the Timeline of immunology:

Jules Alphonse Nicolas Hoffmann is a Luxembourg-born French biologist. During his youth, growing up in Luxembourg, he developed a strong interest in insects under the influence of his father, Jos Hoffmann. This eventually resulted in the younger Hoffmann's dedication to the field of biology using insects as model organisms. He currently holds a faculty position at the University of Strasbourg. He is a research director and member of the board of administrators of the National Center of Scientific Research (CNRS) in Strasbourg, France. He was elected to the positions of Vice-President (2005-2006) and President (2007-2008) of the French Academy of Sciences. Hoffmann and Bruce Beutler were jointly awarded a half share of the 2011 Nobel Prize in Physiology or Medicine for "their discoveries concerning the activation of innate immunity,". [More specifically, the work showing increased Drosomycin expression following activation of Toll pathway in microbial infection.]
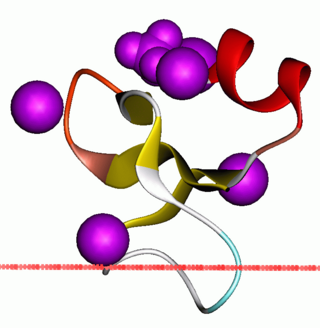
Beta defensins are a family of vertebrate defensins. The beta defensins are antimicrobial peptides implicated in the resistance of epithelial surfaces to microbial colonization.

Beta-defensin 103 is a protein that in humans is encoded by the DEFB103A gene.
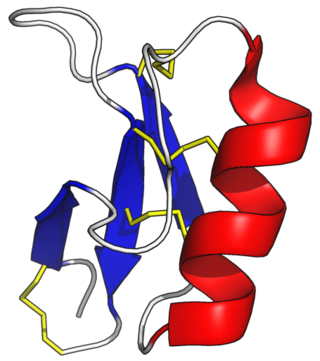
Plant defensins are a family of primitive, highly stable, cysteine-rich defensins found in plants that function to defend them against pathogens and parasites. Defensins are integral components of the innate immune system and belong to the ancient superfamily of antimicrobial peptides (AMPs). AMPs are also known as host defense peptides (HDPs), and they are thought to have diverged about 1.4 billion years ago before the evolution of prokaryotes and eukaryotes. They are ubiquitous in almost all plant species, functionally diverse, and their primary structure varies significantly from one species to the next, except for a few cysteine residues, which stabilize the protein structure through disulfide bond formation. Plant defensins usually have a net positive charge due to the abundance of cationic amino acids and are generally divided into two classes. Those in the class II category contain a C-terminal pro-peptide domain of approximately 33 amino acids and are targeted to the vacuole, while the class I defensins lack this domain and mature in the cell wall. Unlike their class I counterparts, class II plant defensins are relatively smaller, and their acidic C-terminal prodomain is hypothesized to contribute to their vacuolar targeting. The first plant defensins were discovered in barley and wheat in 1990 and were initially designated as γ-thionins. In 1995, the name was changed to 'plant defensin' when it was identified that they are evolutionarily unrelated to other thionins and were more similar to defensins from insects and mammals.

Arthropod defensins are a family defensin proteins found in mollusks, insects, and arachnids. These cysteine-rich antibacterial peptides are primarily active against Gram-positive bacteria and fungi in vitro. However Drosophila fruit flies mutant for the fly defensin were more susceptible to infection by the Gram-negative bacteria Providencia burhodogranariea, and resisted infection against Gram-positive bacteria like wild-type flies. It remains to be seen how in vitro activity relates to in vivo function. Mutants for the defensin-like antimicrobial peptide Drosomycin were more susceptible to fungi, validating a role for defensin-like peptides in anti-fungal defence.
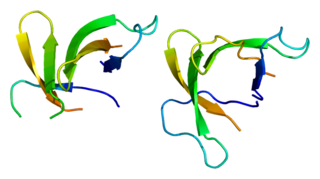
Defensin, alpha 5 (DEFA5) also known as human alpha defensin 5 (HD5) is a protein that in humans is encoded by the DEFA5 gene. DEFA5 is expressed in the Paneth cells of the ileum.

Attacin is a glycine-rich protein of about 20 kDa belonging to the group of antimicrobial peptides (AMP). It is active against Gram-negative bacteria.
Hans Gustaf Boman (1924-2008) was a Swedish microbiologist with a special focus on innate immunity. Boman was born on 16 August 1924 in Engelbrekt Parish, Stockholm, Sweden, and died on 3 December 2008. Boman's pioneering research on insect immunity formed the basis for the Nobel Prize in Physiology or Medicine that was awarded to Jules Hoffman in 2011.

Diptericin is a 9 kDa antimicrobial peptide (AMP) of flies first isolated from the blowfly Phormia terranova. It is primarily active against Gram-negative bacteria, disrupting bacterial membrane integrity. The structure of this protein includes a proline-rich domain with similarities to the AMPs drosocin, pyrrhocoricin, and abaecin, and a glycine-rich domain with similarity to attacin. Diptericin is an iconic readout of immune system activity in flies, used ubiquitously in studies of Drosophila immunity. Diptericin is named after the insect order Diptera.
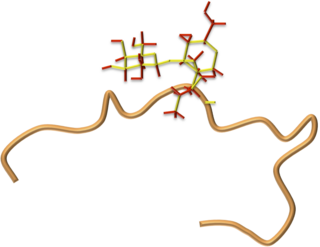
Drosocin is a 19-residue long antimicrobial peptide (AMP) of flies first isolated in the fruit fly Drosophila melanogaster, and later shown to be conserved throughout the genus Drosophila. Drosocin is regulated by the NF-κB Imd signalling pathway in the fly.

Metchnikowin is a 26-residue antimicrobial peptide of the fruit fly Drosophila melanogaster that displays both antibacterial and antifungal properties. This peptide is expressed strongly in the Drosophila fat body, but is also expressed at surface epithelia in the trachea and gut. This is regulated by the NF-κB signalling pathways Toll and Imd. Metchnikowin is named after Russian immunologist Élie Metchnikoff, one of the founders of modern immunology.

The Drosophila quinaria species group is a speciose lineage of mushroom-feeding flies studied for their specialist ecology, their parasites, population genetics, and the evolution of immune systems. Quinaria species are part of the Drosophila subgenus.

The Imd pathway is a broadly-conserved NF-κB immune signalling pathway of insects and some arthropods that regulates a potent antibacterial defence response. The pathway is named after the discovery of a mutation causing severe immune deficiency. The Imd pathway was first discovered in 1995 using Drosophila fruit flies by Bruno Lemaitre and colleagues, who also later discovered that the Drosophila Toll gene regulated defence against Gram-positive bacteria and fungi. Together the Toll and Imd pathways have formed a paradigm of insect immune signalling; as of September 2, 2019, these two landmark discovery papers have been cited collectively over 5000 times since publication on Google Scholar.

Bruno Lemaitre is a French immunologist and a professor at the École Polytechnique Fédérale de Lausanne (EPFL). His research focuses on the mechanisms of innate immunity and endosymbiosis in Drosophila. Lemaitre has also authored several books on the topic of narcissism in science. and a book on the philosophy of Michael Polanyi.
Baramicin (Bara) is an antimicrobial peptide gene of the fruit fly Drosophila melanogaster. Baramicin is a prominent element of the fly immune response: of the most abundant immune peptides detected in the fly hemolymph, the BaraA gene is responsible for 9 of the 24 peptides first described for their high concentrations after systemic infection.
Daisho (Dso) is an antimicrobial peptide gene family of the fruit fly Drosophila melanogaster. Two Daisho genes are encoded in tandem in the fruit fly genome, one shorter than the other. This pair of genes with different length was named "Daisho" in reference to Daisho Japanese swords, which come in pairs with one shorter than the other.
