
Basidiomycota is one of two large divisions that, together with the Ascomycota, constitute the subkingdom Dikarya within the kingdom Fungi. Members are known as basidiomycetes. More specifically, Basidiomycota includes these groups: agarics, puffballs, stinkhorns, bracket fungi, other polypores, jelly fungi, boletes, chanterelles, earth stars, smuts, bunts, rusts, mirror yeasts, and Cryptococcus, the human pathogenic yeast.

Rusts are fungal plant pathogens of the order Pucciniales causing plant fungal diseases.
The hymenium is the tissue layer on the hymenophore of a fungal fruiting body where the cells develop into basidia or asci, which produce spores. In some species all of the cells of the hymenium develop into basidia or asci, while in others some cells develop into sterile cells called cystidia (basidiomycetes) or paraphyses (ascomycetes). Cystidia are often important for microscopic identification. The subhymenium consists of the supportive hyphae from which the cells of the hymenium grow, beneath which is the hymenophoral trama, the hyphae that make up the mass of the hymenophore.

In botany and mycology, a haustorium is a rootlike structure that grows into or around another structure to absorb water or nutrients. For example, in mistletoe or members of the broomrape family, the structure penetrates the host's tissue and draws nutrients from it. In mycology, it refers to the appendage or portion of a parasitic fungus, which performs a similar function. Microscopic haustoria penetrate the host plant's cell wall and siphon nutrients from the space between the cell wall and plasma membrane but do not penetrate the membrane itself. Larger haustoria do this at the tissue level.

A basidiospore is a reproductive spore produced by Basidiomycete fungi, a grouping that includes mushrooms, shelf fungi, rusts, and smuts. Basidiospores typically each contain one haploid nucleus that is the product of meiosis, and they are produced by specialized fungal cells called basidia. Typically, four basidiospores develop on appendages from each basidium, of which two are of one strain and the other two of its opposite strain. In gills under a cap of one common species, there exist millions of basidia. Some gilled mushrooms in the order Agaricales have the ability to release billions of spores. The puffball fungus Calvatia gigantea has been calculated to produce about five trillion basidiospores. Most basidiospores are forcibly discharged, and are thus considered ballistospores. These spores serve as the main air dispersal units for the fungi. The spores are released during periods of high humidity and generally have a night-time or pre-dawn peak concentration in the atmosphere.

Exobasidium vexans is a plant pathogen affecting tea.

Exobasidium vaccinii, commonly known as “red leaf disease,” or “Azalea Gall,” is a biotrophic species of fungus that causes galls on ericaceous plant species, such as blueberry and azalea. Exobasidium vaccinii is considered the type species of the Exobasidium genus. As a member of the Ustilagomycota, it is a basidiomycete closely related to smut fungi. Karl Wilhelm Gottlieb Leopold Fuckel first described the species in 1861 under the basionym Fusidium vaccinii, but in 1867 Mikhail Stepanovich Voronin later placed it in the genus Exobasidium. The type specimen is from Germany, and it is held in the Swedish Museum of Natural History. Exobasidium vaccinii, in current definition from John Axel Nannfeldt in 1981, is limited on the host Vaccinium vitis-idaea. This idea is used in most recent papers on E. vaccinii.

Spongiforma thailandica is a species of fungus in the family Boletaceae, genus Spongiforma. The stemless sponge-like species, first described in 2009, was found in Khao Yai National Park in central Thailand, where it grows in soil in old-growth forests. The rubbery fruit body, which has a strong odor of coal-tar similar to Tricholoma sulphureum, consists of numerous internal cavities lined with spore-producing tissue. Phylogenetic analysis suggests the species is closely related to the Boletaceae genera Porphyrellus and Strobilomyces.

Septobasidium is a fungal genus within the family Septobasidiaceae. Approximately 175 described species are associated with this genus. 227 records are listed by Species Fungorum.

Marasmius sasicola is a species of Marasmiaceae fungus known from Kanagawa Prefecture, Japan. First collected in 2000, it was described in 2002 by Haruki Takahashi. The species produces small mushrooms with white caps and very short, very thin black stems. Unlike in other, similar species, the stems enter the plant matter on which the mushroom grows. The six to eight white gills are spread out around the cap, and all of them reach the stem. The flesh has no taste or odour. Found in June, the species grows on dead Sasa leaves, from which it takes its specific epithet.

Auriscalpium vulgare, commonly known as the pinecone mushroom, the cone tooth, or the ear-pick fungus, is a species of fungus in the family Auriscalpiaceae of the order Russulales. It was first described in 1753 by Carl Linnaeus, who included it as a member of the tooth fungi genus Hydnum, but British mycologist Samuel Frederick Gray recognized its uniqueness and in 1821 transferred it to the genus Auriscalpium that he created to contain it. The fungus is widely distributed in Europe, Central America, North America, and temperate Asia. Although common, its small size and nondescript colors lead it to be easily overlooked in the pine woods where it grows. A. vulgare is not generally considered edible because of its tough texture, but some historical literature says it used to be consumed in France and Italy.
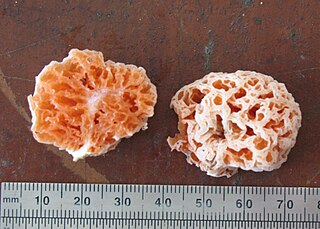
Spongiforma is a genus of sponge-like fungi in the family Boletaceae. Newly described in 2009, the genus contains two species: S. thailandica and S. squarepantsii. The type species S. thailandica is known only from Khao Yai National Park in central Thailand, where it grows in soil in old-growth forests dominated by dipterocarp trees. The rubbery fruit bodies, which has a strong odour of coal-tar similar to Tricholoma sulphureum, consists of numerous internal cavities lined with spore-producing tissue. S. squarepantsii, described as new to science in 2011, is found in Malaysia. It produces sponge-like, rubbery orange fruit bodies with a fruity or musky odour. These fruit bodies will—like a sponge—resume their original shape if water is squeezed out. The origin of the specific name derives from its perceived resemblance to the cartoon character SpongeBob SquarePants. Apart from differences in distribution, S. squarepantsii differs from S. thailandica in its colour, odour, and spore structure.

Myxarium nucleatum is a species of fungus in the family Hyaloriaceae. In the UK, it has been given the recommended English name of crystal brain. Basidiocarps are watery white, pustular or lobed, and gelatinous with small, white, mineral inclusions visible to the naked eye. It is a common, wood-rotting species in Europe, typically growing on dead attached or fallen branches of broadleaf trees. It is currently not clear whether collections from North America and elsewhere represent the same species.

Phellinus ellipsoideus is a species of polypore fungus in the family Hymenochaetaceae, a specimen of which produced the largest fungal fruit body ever recorded. Found in China, the fruit bodies produced by the species are brown, woody basidiocarps that grow on dead wood, where the fungus feeds as a saprotroph. The basidiocarps are perennial, allowing them to grow very large under favourable circumstances. They are resupinate, measuring 30 centimetres (12 in) or more in length, though typically extending less than a centimetre from the surface of the wood. P. ellipsoideus produces distinct ellipsoidal spores, after which it is named, and unusual setae. These two features allow it to be readily differentiated microscopically from other, similar species. Chemical compounds isolated from the species include several steroidal compounds. These may have pharmacological applications, but further research is needed.

Phaeotremella frondosa is a species of fungus in the family Phaeotremellaceae producing brownish, frondose, gelatinous basidiocarps. It is widespread in north temperate regions, and is parasitic on other species of fungi that grow on dead attached and recently fallen branches of broadleaf trees.

Naematelia aurantia is a species of fungus producing yellow, frondose, gelatinous basidiocarps. It is widespread in north temperate regions and is parasitic on another species of fungus that grows on dead attached and recently fallen branches of broadleaf trees. It is commonly called golden ear in North America.
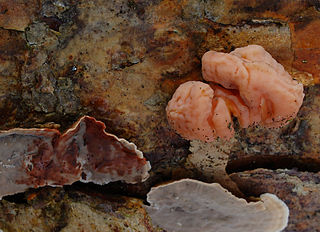
Naematelia encephala is a species of fungus producing pink, brain-like, gelatinous basidiocarps. It is widespread in north temperate regions and is parasitic on another species of fungus that grows on dead attached and recently fallen branches of conifers. In the UK, its recommended English name is conifer brain.
The Melaniellaceae are a family of fungi in the division Basidiomycota and order of Doassansiales. The family contains 1 genera and 2 species. They have a distribution in south and south-east Asia.
Zygogloea is a genus of fungi in the phylum Basidiomycota. The type and only species, Zygogloea gemellipara, is an hymenial parasite of Myxarium nucleatum and produces no visible basidiocarps of its own. The species has "auricularioid" basidia, septate basidiospores, and forms unusual twinned zygoconidia. The hyphae of the parasite attach to the host hyphae through twisting, tendril-like haustorial cells. Zygogloea gemellipara was described from England and has also been reported from Belgium. The genus has been referred to the Pucciniomycotina, but its place within this subdivision is unknown.
Tremella anaptychiae is a species of lichenicolous (lichen-dwelling) fungus in the family Tremellaceae. It was first reported in the literature in 1996 by mycologist Paul Diederich, who did not formally describe it as a new species due to the paucity of material. Additional material was collected in later years, and it was finally described in 2017 by Juan Carlos Zamora and Diederich. The fungus is known to occur in Italy, Macedonia, Spain, and Sweden. It is confined to the host lichen Anaptychia ciliaris, which has a largely palearctic distribution.
