Related Research Articles

An ecosystem consists of all the organisms and the physical environment with which they interact. These biotic and abiotic components are linked together through nutrient cycles and energy flows. Energy enters the system through photosynthesis and is incorporated into plant tissue. By feeding on plants and on one another, animals play an important role in the movement of matter and energy through the system. They also influence the quantity of plant and microbial biomass present. By breaking down dead organic matter, decomposers release carbon back to the atmosphere and facilitate nutrient cycling by converting nutrients stored in dead biomass back to a form that can be readily used by plants and microbes.

A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, its root system. Mycorrhizae play important roles in plant nutrition, soil biology, and soil chemistry.
This glossary of ecology is a list of definitions of terms and concepts in ecology and related fields. For more specific definitions from other glossaries related to ecology, see Glossary of biology, Glossary of evolutionary biology, and Glossary of environmental science.

An arbuscular mycorrhiza (AM) is a type of mycorrhiza in which the symbiont fungus penetrates the cortical cells of the roots of a vascular plant forming arbuscules. Arbuscular mycorrhiza is a type of endomycorrhiza along with ericoid mycorrhiza and orchid mycorrhiza.

Biological soil crusts are communities of living organisms on the soil surface in arid and semi-arid ecosystems. They are found throughout the world with varying species composition and cover depending on topography, soil characteristics, climate, plant community, microhabitats, and disturbance regimes. Biological soil crusts perform important ecological roles including carbon fixation, nitrogen fixation and soil stabilization; they alter soil albedo and water relations and affect germination and nutrient levels in vascular plants. They can be damaged by fire, recreational activity, grazing and other disturbances and can require long time periods to recover composition and function. Biological soil crusts are also known as biocrusts or as cryptogamic, microbiotic, microphytic, or cryptobiotic soils.
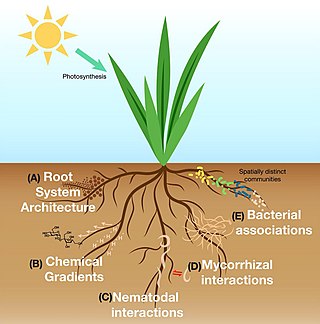
The rhizosphere is the narrow region of soil or substrate that is directly influenced by root secretions and associated soil microorganisms known as the root microbiome. Soil pores in the rhizosphere can contain many bacteria and other microorganisms that feed on sloughed-off plant cells, termed rhizodeposition, and the proteins and sugars released by roots, termed root exudates. This symbiosis leads to more complex interactions, influencing plant growth and competition for resources. Much of the nutrient cycling and disease suppression by antibiotics required by plants, occurs immediately adjacent to roots due to root exudates and metabolic products of symbiotic and pathogenic communities of microorganisms. The rhizosphere also provides space to produce allelochemicals to control neighbours and relatives.

Soil biology is the study of microbial and faunal activity and ecology in soil. Soil life, soil biota, soil fauna, or edaphon is a collective term that encompasses all organisms that spend a significant portion of their life cycle within a soil profile, or at the soil-litter interface. These organisms include earthworms, nematodes, protozoa, fungi, bacteria, different arthropods, as well as some reptiles, and species of burrowing mammals like gophers, moles and prairie dogs. Soil biology plays a vital role in determining many soil characteristics. The decomposition of organic matter by soil organisms has an immense influence on soil fertility, plant growth, soil structure, and carbon storage. As a relatively new science, much remains unknown about soil biology and its effect on soil ecosystems.
Soil ecology is the study of the interactions among soil organisms, and between biotic and abiotic aspects of the soil environment. It is particularly concerned with the cycling of nutrients, formation and stabilization of the pore structure, the spread and vitality of pathogens, and the biodiversity of this rich biological community.

Soil respiration refers to the production of carbon dioxide when soil organisms respire. This includes respiration of plant roots, the rhizosphere, microbes and fauna.

Mycoforestry is an ecological forest management system implemented to enhance forest ecosystems and plant communities through the introduction of mycorrhizal and saprotrophic fungi. Mycoforestry is considered a type of permaculture and can be implemented as a beneficial component of an agroforestry system. Mycoforestry can enhance the yields of tree crops and produce edible mushrooms, an economically valuable product. By integrating plant-fungal associations into a forestry management system, native forests can be preserved, wood waste can be recycled back into the ecosystem, carbon sequestration can be increased, planted restoration sites are enhanced, and the sustainability of forest ecosystems are improved. Mycoforestry is an alternative to the practice of clearcutting, which removes dead wood from forests, thereby diminishing nutrient availability and reducing soil depth.

A mycorrhizal network is an underground network found in forests and other plant communities, created by the hyphae of mycorrhizal fungi joining with plant roots. This network connects individual plants together. Mycorrhizal relationships are most commonly mutualistic, with both partners benefiting, but can be commensal or parasitic, and a single partnership may change between any of the three types of symbiosis at different times.

Soil carbon storage is an important function of terrestrial ecosystems. Soil contains more carbon than plants and the atmosphere combined. Understanding what maintains the soil carbon pool is important to understand the current distribution of carbon on Earth, and how it will respond to environmental change. While much research has been done on how plants, free-living microbial decomposers, and soil minerals affect this pool of carbon, it is recently coming to light that mycorrhizal fungi—symbiotic fungi that associate with roots of almost all living plants—may play an important role in maintaining this pool as well. Measurements of plant carbon allocation to mycorrhizal fungi have been estimated to be 5 to 20% of total plant carbon uptake, and in some ecosystems the biomass of mycorrhizal fungi can be comparable to the biomass of fine roots. Recent research has shown that mycorrhizal fungi hold 50 to 70 percent of the total carbon stored in leaf litter and soil on forested islands in Sweden. Turnover of mycorrhizal biomass into the soil carbon pool is thought to be rapid and has been shown in some ecosystems to be the dominant pathway by which living carbon enters the soil carbon pool.

Extracellular enzymes or exoenzymes are synthesized inside the cell and then secreted outside the cell, where their function is to break down complex macromolecules into smaller units to be taken up by the cell for growth and assimilation. These enzymes degrade complex organic matter such as cellulose and hemicellulose into simple sugars that enzyme-producing organisms use as a source of carbon, energy, and nutrients. Grouped as hydrolases, lyases, oxidoreductases and transferases, these extracellular enzymes control soil enzyme activity through efficient degradation of biopolymers.

An ectomycorrhiza is a form of symbiotic relationship that occurs between a fungal symbiont, or mycobiont, and the roots of various plant species. The mycobiont is often from the phyla Basidiomycota and Ascomycota, and more rarely from the Zygomycota. Ectomycorrhizas form on the roots of around 2% of plant species, usually woody plants, including species from the birch, dipterocarp, myrtle, beech, willow, pine and rose families. Research on ectomycorrhizas is increasingly important in areas such as ecosystem management and restoration, forestry and agriculture.

Ectomycorrhizal extramatrical mycelium is the collection of filamentous fungal hyphae emanating from ectomycorrhizas. It may be composed of fine, hydrophilic hypha which branches frequently to explore and exploit the soil matrix or may aggregate to form rhizomorphs; highly differentiated, hydrophobic, enduring, transport structures.

The root microbiome is the dynamic community of microorganisms associated with plant roots. Because they are rich in a variety of carbon compounds, plant roots provide unique environments for a diverse assemblage of soil microorganisms, including bacteria, fungi and archaea. The microbial communities inside the root and in the rhizosphere are distinct from each other, and from the microbial communities of bulk soil, although there is some overlap in species composition.

Cenococcum geophilum Fr., synonym Cenococcum graniforme (Sow.) Ferd. and Winge, is an Ascomycete fungal species and is the only member in the genus Cenococcum. It is one of the most common ectomycorrhizal fungal species encountered in forest ecosystems. The geographic distribution of the species is notably cosmopolitan; it is found in ecosystems with a wide range of environmental conditions, and in many cases in high relative frequency. Because of its wide distribution and abundance in forest soils, it is one of the most well-studied ectomycorrhizal fungal species. While the species has long been known to be sterile and not produce asexual or sexual spores, cryptic sexual stages may exist. The hyphae produced by C. geophilum are characterized by their thick (1.5-8 um), straight and jet black appearance with little branching. They usually form monopodial (unbranched) ectomycorrhizas. The mantles of C. geophilum ectomycorrhizas are usually thick with few to many emanating hyphae.
Dark septate endophytes (DSE) are a group of endophytic fungi characterized by their morphology of melanized, septate, hyphae. This group is likely paraphyletic, and contain conidial as well as sterile fungi that colonize roots intracellularly or intercellularly. Very little is known about the number of fungal taxa within this group, but all are in the Ascomycota. They are found in over 600 plant species and across 114 families of angiosperms and gymnosperms and co-occur with other types of mycorrhizal fungi. They have a wide global distribution and can be more abundant in stressed environments. Much of their taxonomy, physiology, and ecology are unknown.
Orchid mycorrhizae are endomycorrhizal fungi which develop symbiotic relationships with the roots and seeds of plants of the family Orchidaceae. Nearly all orchids are myco-heterotrophic at some point in their life cycle. Orchid mycorrhizae are critically important during orchid germination, as an orchid seed has virtually no energy reserve and obtains its carbon from the fungal symbiont.
Saprotrophic bacteria are bacteria that are typically soil-dwelling and utilize saprotrophic nutrition as their primary energy source. They are often associated with soil fungi that also use saprotrophic nutrition and both are classified as saprotrophs.
References
- 1 2 3 Collins, S.L., Sinsabaugh, R.L., Crenshaw, C., Green, L., Porras-Alfaro, A., Stursova, M., and Zeglin, L.H. 2008. Pulse dynamics and microbial processes in aridland ecosystems. Journal of Ecology 96: 413–420.
- ↑ Williams, C.A., Hanan, N., Scholes, R.J., Kutsch, W. 2009. Complexity in water and carbon dioxide fluxes following rain pulses in an African savanna. Oecologia 161: 469-480.
- ↑ Charley, J.L., and West, N.E. 1975. Plant-Induced Soil Chemical Patterns in Some Shrub-Dominated Semi-Desert Ecosystems of Utah. Journal of Ecology 63(3): 945-963.
- ↑ Schlesinger, W.H., and Pilmanis, A.M. 1998. Plant-soil interactions in deserts. Biogeochemistry 42: 169-187.
- ↑ Hawkes, C. 2003. Nitrogen cycling mediated by biological soil crusts and arbuscular mycorrhizal fungi. Ecology 84: 1553-1562.
- ↑ Skujins, R.G. 1976. Extracellular enzymes in soil. CRC Critical Reviews in Microbiology. 4(4): 383-421.
- ↑ Allen, M.F. 2007. Mycorrhizal Fungi: Highways for Water and Nutrients in Arid Soils. Vadose Zone Journal 6(2): 291-297.
- ↑ Mandyam, K. and Jumpponen, A. 2005. Seeking the elusive function of the root-colonising dark septate endophytic fungi. Studies in Mycology. 53: 173-189.
- ↑ Porras-Alfaro, A., Herrera, J., Natvig, D.O., Lipinski, K., Sinsabaugh, R.L. 2011. Diversity and distribution of soil fungal communities in a semiarid grassland. Mycologia 103: 10-21.
- ↑ Green, L.E., Porras-Alfaro, A., and Sinsabaugh, R.L. 2008. Translocation of nitrogen and carbon integrates biotic crust and grass production in desert grassland. Journal of Ecology 96: 1076–1085.
- ↑ Allen, M. F. (2009), Bidirectional water flows through the soil–fungal–plant mycorrhizal continuum. New Phytologist, 182: 290–293.