
Rusts are fungal plant pathogens of the order Pucciniales causing plant fungal diseases.

Stem rust, also known as cereal rust, black rust, red rust or red dust, is caused by the fungus Puccinia graminis, which causes significant disease in cereal crops. Crop species that are affected by the disease include bread wheat, durum wheat, barley and triticale. These diseases have affected cereal farming throughout history. The annual recurrence of stem rust of wheat in North Indian plains was discovered by K.C. Mehta. Since the 1950s, wheat strains bred to be resistant to stem rust have become available. Fungicides effective against stem rust are available as well.

Venturia inaequalis is an ascomycete fungus that causes the apple scab disease.

Hypersensitive response (HR) is a mechanism used by plants to prevent the spread of infection by microbial pathogens. HR is characterized by the rapid death of cells in the local region surrounding an infection and it serves to restrict the growth and spread of pathogens to other parts of the plant. It is analogous to the innate immune system found in animals, and commonly precedes a slower systemic response, which ultimately leads to systemic acquired resistance (SAR). HR can be observed in the vast majority of plant species and is induced by a wide range of plant pathogens such as oomycetes, viruses, fungi and even insects.
The gene-for-gene relationship is a concept in plant pathology that plants and their diseases each have single genes that interact with each other during an infection. It was proposed by Harold Henry Flor who was working with rust (Melampsora lini) of flax (Linum usitatissimum). Flor showed that the inheritance of both resistance in the host and parasite ability to cause disease is controlled by pairs of matching genes. One is a plant gene called the resistance (R) gene. The other is a parasite gene called the avirulence (Avr) gene. Plants producing a specific R gene product are resistant towards a pathogen that produces the corresponding Avr gene product. Gene-for-gene relationships are a widespread and very important aspect of plant disease resistance. Another example can be seen with Lactuca serriola versus Bremia lactucae.
Leptosphaeria maculans is a fungal pathogen of the phylum Ascomycota that is the causal agent of blackleg disease on Brassica crops. Its genome has been sequenced, and L. maculans is a well-studied model phytopathogenic fungus. Symptoms of blackleg generally include basal stem cankers, small grey lesions on leaves, and root rot. The major yield loss is due to stem canker. The fungus is dispersed by the wind as ascospores or rain splash in the case of the conidia. L. maculans grows best in wet conditions and a temperature range of 5–20 degrees Celsius. Rotation of crops, removal of stubble, application of fungicide, and crop resistance are all used to manage blackleg. The fungus is an important pathogen of Brassica napus (canola) crops.
This is a glossary of some of the terms used in phytopathology.

Xanthomonas campestris is a gram-negative, obligate aerobic bacterium that is a member of the Xanthomonas genus, which is a group of bacteria that are commonly known for their association with plant disease. This species includes Xanthomonas campestris pv. campestris the cause of black rot of brassicas, one of the most important diseases of brasicas worldwide.

Pseudomonas syringae is a rod-shaped, Gram-negative bacterium with polar flagella. As a plant pathogen, it can infect a wide range of species, and exists as over 50 different pathovars, all of which are available to researchers from international culture collections such as the NCPPB, ICMP, and others.
Ug99 is a lineage of wheat stem rust, which is present in wheat fields in several countries in Africa and the Middle East and is predicted to spread rapidly through these regions and possibly further afield, potentially causing a wheat production disaster that would affect food security worldwide. In 2005 the noted green revolution pioneer Norman Borlaug brought great attention to the problem, and most subsequent efforts can be traced to his advocacy. It can cause up to 100% crop losses and is virulent against many resistance genes which have previously protected wheat against stem rust.

Melampsora lini is a species of fungus and plant pathogen found in Ireland and commonly known as flax rust.

Melampsora medusae is a fungal pathogen, causing a disease of woody plants. The infected trees' leaves turn yellowish-orange. The disease affects mostly conifers, e.g. the Douglas-fir, western larch, tamarack, ponderosa, and lodgepole pine trees, but also some broadleaves, e.g. trembling aspen and poplars. Coniferous hosts are affected in late spring through early August, and trembling aspens and poplars from early summer to late fall. It is one of only two foliage rusts that occur naturally in British Columbia.

Linum catharticum, also known as purging flax, or fairy flax, is an herbaceous flowering plant in the family Linaceae, native to Great Britain, Iceland, central Europe and Western Asia. It is an annual plant and blooms in July and August.
Resistance genes (R-Genes) are genes in plant genomes that convey plant disease resistance against pathogens by producing R proteins. The main class of R-genes consist of a nucleotide binding domain (NB) and a leucine rich repeat (LRR) domain(s) and are often referred to as (NB-LRR) R-genes or NLRs. Generally, the NB domain binds either ATP/ADP or GTP/GDP. The LRR domain is often involved in protein-protein interactions as well as ligand binding. NB-LRR R-genes can be further subdivided into toll interleukin 1 receptor (TIR-NB-LRR) and coiled-coil (CC-NB-LRR).

Plant disease resistance protects plants from pathogens in two ways: by pre-formed structures and chemicals, and by infection-induced responses of the immune system. Relative to a susceptible plant, disease resistance is the reduction of pathogen growth on or in the plant, while the term disease tolerance describes plants that exhibit little disease damage despite substantial pathogen levels. Disease outcome is determined by the three-way interaction of the pathogen, the plant and the environmental conditions.
Host–parasite coevolution is a special case of coevolution, where a host and a parasite continually adapt to each other. This can create an evolutionary arms race between them. A more benign possibility is of an evolutionary trade-off between transmission and virulence in the parasite, as if it kills its host too quickly, the parasite will not be able to reproduce either. Another theory, the Red Queen hypothesis, proposes that since both host and parasite have to keep on evolving to keep up with each other, and since sexual reproduction continually creates new combinations of genes, parasitism favours sexual reproduction in the host.

Harold Henry Flor known as H. H. Flor (1900–1991) was an American plant pathologist famous for proposing the gene for gene hypothesis of plant-pathogen genetic interaction whilst working on rust of flax.
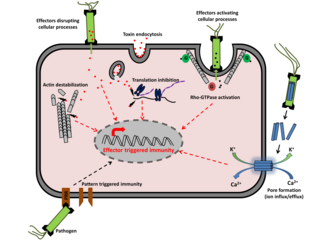
Effector-triggered immunity (ETI) is one of the pathways, along with the pattern-triggered immunity (PTI) pathway, by which the innate immune system recognises pathogenic organisms and elicits a protective immune response. ETI is elicited when an effector protein secreted by a pathogen into the host cell is successfully recognised by the host. Alternatively, effector-triggered susceptibility (ETS) can occur if an effector protein can block the immune response triggered by pattern recognition receptors (PRR) and evade immunity, allowing the pathogen to propagate in the host.

Bacterial blight of soybean is a widespread disease of soybeans caused by Pseudomonas syringaepv. glycinea.
Martin Parniske is a German biologist with a specialisation in genetics, microbiology and biochemistry. He is university professor and head of the Institute of Genetics at the Faculty of Biology of the Ludwig Maximilian University of Munich. Parniske's scientific focus is on the molecular interaction between plants and symbiotic and pathogenic organisms including bacteria, fungi, oomycetes and insects.
