
The lophophore is a characteristic feeding organ possessed by four major groups of animals: the Brachiopoda, Bryozoa, Hyolitha, and Phoronida, which collectively constitute the protostome group Lophophorata. All lophophores are found in aquatic organisms.

The halkieriids are a group of fossil organisms from the Lower to Middle Cambrian. Their eponymous genus is Halkieria, which has been found on almost every continent in Lower to Mid Cambrian deposits, forming a large component of the small shelly fossil assemblages. The best known species is Halkieria evangelista, from the North Greenland Sirius Passet Lagerstätte, in which complete specimens were collected on an expedition in 1989. The fossils were described by Simon Conway Morris and John Peel in a short paper in 1990 in the journal Nature. Later a more thorough description was undertaken in 1995 in the journal Philosophical Transactions of the Royal Society of London and wider evolutionary implications were posed.
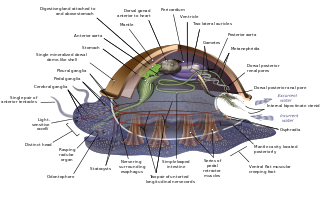
The evolution of the molluscs is the way in which the Mollusca, one of the largest groups of invertebrate animals, evolved. This phylum includes gastropods, bivalves, scaphopods, cephalopods, and several other groups. The fossil record of mollusks is relatively complete, and they are well represented in most fossil-bearing marine strata. Very early organisms which have dubiously been compared to molluscs include Kimberella and Odontogriphus.

Haplophrentis is a genus of tiny shelled hyolithid which lived in the Cambrian Period. Its shell was long and conical, with the open end protected by an operculum, from which two fleshy arms called helens protruded at the sides. These arms served to elevate the opening of the shells above the sea floor, acting like stilts.

Yochelcionella is an extinct genus of basal molluscs which lived during the Tommotian epoch, the first epoch of the Cambrian period. This genus is often reconstructed to resemble snails.

Helcionellid or Helcionelliformes is an order of small fossil shells that are universally interpreted as molluscs, though no sources spell out why this taxonomic interpretation is preferred. These animals are first found about 540 to 530 million years ago in the late Nemakit-Daldynian age, which is the earliest part of the Cambrian period. A single species persisted to the Early Ordovician. These fossils are component of the small shelly fossils (SSF) assemblages.

Phoronids are a small phylum of marine animals that filter-feed with a lophophore, and build upright tubes of chitin to support and protect their soft bodies. They live in most of the oceans and seas, including the Arctic Ocean but excluding the Antarctic Ocean, and between the intertidal zone and about 400 meters down. Most adult phoronids are 2 cm long and about 1.5 mm wide, although the largest are 50 cm long.
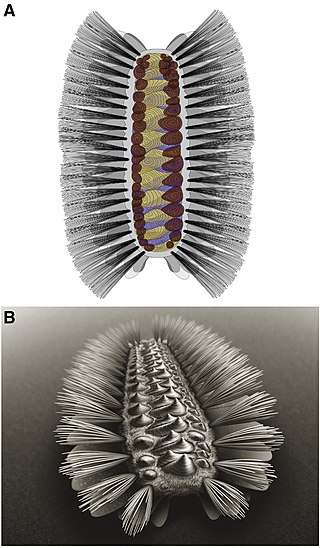
Tommotiids are an extinct group of Cambrian invertebrates thought to be early lophophorates.

The origin of the brachiopods is uncertain; they either arose from reduction of a multi-plated tubular organism, or from the folding of a slug-like organism with a protective shell on either end. Since their Cambrian origin, the phylum rose to a Palaeozoic dominance, but dwindled during the Mesozoic.
Stenothecidae is an extinct family of fossil univalved Cambrian molluscs which may be either gastropods or monoplacophorans.
Anabarella is a species of bilaterally-flattened monoplacophoran mollusc, with a morphological similarity to the rostroconchs. Its shell preserves evidence of three mineralogical textures on its outer surface: it is polygonal near the crest of the shell, subsequently changing to both spiny and stepwise. Its internal microstructure is calcitic and semi-nacreous. Its name reflects its provenance from Anabar, Siberia. It has been interpreted as ancestral to the rostroconchs, and has been aligned to the Helcionellidae.

Fordilla is an extinct genus of early bivalves, one of two genera in the extinct family Fordillidae. The genus is known solely from Early Cambrian fossils found in North America, Greenland, Europe, the Middle East, and Asia. The genus currently contains three described species, Fordilla germanica, Fordilla sibirica, and the type species Fordilla troyensis.
The cephalopods have a long geological history, with the first nautiloids found in late Cambrian strata, and purported stem-group representatives present in the earliest Cambrian lagerstätten.
Stenothecoida is a taxon of bivalved fossils from the Early to middle Cambrian period. They look a bit like brachiopods or bivalve molluscs.
Watsonella is an extinct genus of mollusc known from early (Terreneuvian) Cambrian strata. It has been hypothesized to be close to the origin of bivalves. It contains a single species, Watsonella crosbyi.
The orthothecids are one of the two hyolith orders.

The Hyolithida are lophophorates, one of the two orders of hyolithid, the other being the Orthothecida. Most of our knowledge of the hyolithids comes from studies on the Hyolithida. Both orders had an operculum that was not hinged to the conch. However, the Hyolithida are distinct from the Orthothecida in having additional paired, curved, whiskerlike appendages. The Hyolithida were probably bottom feeders living in shallow water, and had tentacules.
Hyolithellus is a conical tubular fossil from the Cambrian, originally considered a hyolith but since reinterpreted tentatively as an annelid worm.
Cupitheca is a genus of Cambrian hyolith with the unusual distinction of shedding the apex of its camerate conical shell. As with Triplicatella and Hyptiotheca, its designation to the hyolithids or orthothecids is not straightforward, exhibiting as it does a mixture of the characters that would normally demark the two subtaxa of Hyolitha.



