
Tsetse are large, biting flies that inhabit much of tropical Africa. Tsetse flies include all the species in the genus Glossina, which are placed in their own family, Glossinidae. The tsetse is an obligate parasite, which lives by feeding on the blood of vertebrate animals. Tsetse has been extensively studied because of their role in transmitting disease. They have a pronounced economic impact in sub-Saharan Africa as the biological vectors of trypanosomes, causing human and animal trypanosomiasis.

Trypanosomiasis or trypanosomosis is the name of several diseases in vertebrates caused by parasitic protozoan trypanosomes of the genus Trypanosoma. In humans this includes African trypanosomiasis and Chagas disease. A number of other diseases occur in other animals.

Vector control is any method to limit or eradicate the mammals, birds, insects or other arthropods which transmit disease pathogens. The most frequent type of vector control is mosquito control using a variety of strategies. Several of the "neglected tropical diseases" are spread by such vectors.

Parotitis is an inflammation of one or both parotid glands, the major salivary glands located on either side of the face, in humans. The parotid gland is the salivary gland most commonly affected by inflammation.

The adaptive immune system, also known as the acquired immune system, or specific immune system is a subsystem of the immune system that is composed of specialized, systemic cells and processes that eliminate pathogens or prevent their growth. The acquired immune system is one of the two main immunity strategies found in vertebrates.
White spot syndrome (WSS) is a viral infection of penaeid shrimp. The disease is highly lethal and contagious, killing shrimp quickly. Outbreaks of this disease have wiped out the entire populations of many shrimp farms within a few days, in places throughout the world.
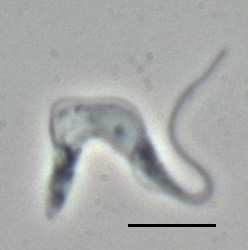
Trypanosoma brucei is a species of parasitic kinetoplastid belonging to the genus Trypanosoma that is present in sub-Saharan Africa. Unlike other protozoan parasites that normally infect blood and tissue cells, it is exclusively extracellular and inhabits the blood plasma and body fluids. It causes deadly vector-borne diseases: African trypanosomiasis or sleeping sickness in humans, and animal trypanosomiasis or nagana in cattle and horses. It is a species complex grouped into three subspecies: T. b. brucei, T. b. gambiense and T. b. rhodesiense. The first is a parasite of non-human mammals and causes nagana, while the latter two are zoonotic infecting both humans and animals and cause African trypanosomiasis.

Animal trypanosomiasis, also known as nagana and nagana pest, or sleeping sickness, is a disease of vertebrates. The disease is caused by trypanosomes of several species in the genus Trypanosoma such as T. brucei. T. vivax causes nagana mainly in West Africa, although it has spread to South America. The trypanosomes infect the blood of the vertebrate host, causing fever, weakness, and lethargy, which lead to weight loss and anemia; in some animals the disease is fatal unless treated. The trypanosomes are transmitted by tsetse flies.

Murine coronavirus (M-CoV) is a virus in the genus Betacoronavirus that infects mice. Belonging to the subgenus Embecovirus, murine coronavirus strains are enterotropic or polytropic. Enterotropic strains include mouse hepatitis virus (MHV) strains D, Y, RI, and DVIM, whereas polytropic strains, such as JHM and A59, primarily cause hepatitis, enteritis, and encephalitis. Murine coronavirus is an important pathogen in the laboratory mouse and the laboratory rat. It is the most studied coronavirus in animals other than humans, and has been used as an animal disease model for many virological and clinical studies.
Paratransgenesis is a technique that attempts to eliminate a pathogen from vector populations through transgenesis of a symbiont of the vector. The goal of this technique is to control vector-borne diseases. The first step is to identify proteins that prevent the vector species from transmitting the pathogen. The genes coding for these proteins are then introduced into the symbiont, so that they can be expressed in the vector. The final step in the strategy is to introduce these transgenic symbionts into vector populations in the wild. One use of this technique is to prevent mortality for humans from insect-borne diseases. Preventive methods and current controls against vector-borne diseases depend on insecticides, even though some mosquito breeds may be resistant to them. There are other ways to fully eliminate them. “Paratransgenesis focuses on utilizing genetically modified insect symbionts to express molecules within the vector that are deleterious to pathogens they transmit.” The acidic bacteria Asaia symbionts are beneficial in the normal development of mosquito larvae; however, it is unknown what Asais symbionts do to adult mosquitoes.

The desert warthog is a species of even-toed ungulate in the pig family (Suidae), found in northern Kenya and Somalia, and possibly Djibouti, Eritrea, and Ethiopia. This is the range of the extant subspecies, commonly known as the Somali warthog. Another subspecies, commonly known as the Cape warthog, became extinct around 1865, but formerly occurred in South Africa.

The housefly is a fly of the suborder Cyclorrhapha. It possibly originated in the Middle East, and spread around the world as a commensal of humans. It is the most common fly species found in houses. Adults are gray to black, with four dark, longitudinal lines on the thorax, slightly hairy bodies, and a single pair of membranous wings. They have red eyes, set farther apart in the slightly larger female.

Salivary gland diseases (SGDs) are multiple and varied in cause. There are three paired major salivary glands in humans: the parotid glands, the submandibular glands, and the sublingual glands. There are also about 800–1,000 minor salivary glands in the mucosa of the mouth. The parotid glands are in front of the ears, one on side, and secrete mostly serous saliva, via the parotid ducts, into the mouth, usually opening roughly opposite the second upper molars. The submandibular gland is medial to the angle of the mandible, and it drains its mixture of serous and mucous saliva via the submandibular duct into the mouth, usually opening in a punctum in the floor of mouth. The sublingual gland is below the tongue, on the floor of the mouth; it drains its mostly mucous saliva into the mouth via about 8–20 ducts, which open along the plica sublingualis, a fold of tissue under the tongue.
Glossinavirus is a genus of viruses, in the family Hytrosaviridae. Glossina sp serve as natural hosts. There is only one species in this genus: Glossina hytrosavirus. Diseases associated with this genus include: partial sterility due to ovarian abnormalities or to testicular degeneration; can be asymptomatic in laboratory colonies.
Muscavirus is a genus of viruses, in the family Hytrosaviridae. The fly Musca domestica is the natural host. There is only one species in this genus: Musca hytrosavirus. Diseases associated with this genus include: salivary gland hypertrophy, and complete sterility of infected female flies by inhibiting eggs development.
Glossina fuscipes is a riverine fly species in the genus Glossina, which are commonly known as tsetse flies. Typically found in sub-Saharan Africa but with a small Arabian range, G. fuscipes is a regional vector of African trypanosomiasis, commonly known as sleeping sickness, that causes significant rates of morbidity and mortality among humans and livestock. Consequently, the species is among several being targeted by researchers for population control as a method for controlling the disease.
Rat coronavirus, or RCV, is a strain or subspecies of Murine coronavirus that infects rats. The earliest discovered strains of Rat coronavirus were Sialodacryoadenitis virus, also known as SDAV, and Parker's Rat Coronavirus (PRC). Four other substrains have since been discovered; CARS, RCV-BCMM, RCV-W and RCV-NJ.

Glossina morsitans is a species of tsetse fly in the genus Glossina. It is one of the major vectors of Trypanosoma brucei rhodesiense in African savannas.
The Sleeping Sickness Commission was a medical project established by the British Royal Society to investigate the outbreak of African sleeping sickness or African trypanosomiasis in Africa at the turn of the 20th century. The outbreak of the disease started in 1900 in Uganda, which was at the time a protectorate of the British Empire. The initial team in 1902 consisted of Aldo Castellani and George Carmichael Low, both from the London School of Hygiene and Tropical Medicine, and Cuthbert Christy, a medical officer on duty in Bombay, India. From 1903, David Bruce of the Royal Army Medical Corps and David Nunes Nabarro of the University College Hospital took over the leadership. The commission established that species of blood protozoan called Trypanosoma brucei, named after Bruce, was the causative parasite of sleeping sickness.
Naldaviricetes is a class of viruses, which infect arthropods. Members of Naldaviricetes are characterized by large enveloped rod-shaped virions, circular double-stranded DNA genomes, and replication in the nucleus of the host cell. All of them share a set of unique genes not found in other viruses, which include the presence of multiple interspersed direct repeats, various subunits of DNA polymerase and RNA polymerase, four late expression factor genes, and infectivity factor genes suggesting a common host entry mechanism.