Vertebrates are deuterostomal animals with bony or cartilaginous axial endoskeleton — known as the vertebral column, spine or backbone — around and along the spinal cord, including all fish, amphibians, reptiles, birds and mammals. The vertebrates consist of all the taxa within the subphylum Vertebrata and represent the overwhelming majority of the phylum Chordata, with currently about 69,963 species described.

Inbreeding is the production of offspring from the mating or breeding of individuals or organisms that are closely related genetically. By analogy, the term is used in human reproduction, but more commonly refers to the genetic disorders and other consequences that may arise from expression of deleterious recessive traits resulting from incestuous sexual relationships and consanguinity. Animals avoid incest only rarely.

Behavioral ecology, also spelled behavioural ecology, is the study of the evolutionary basis for animal behavior due to ecological pressures. Behavioral ecology emerged from ethology after Niko Tinbergen outlined four questions to address when studying animal behaviors: What are the proximate causes, ontogeny, survival value, and phylogeny of a behavior?

Sperm competition is the competitive process between spermatozoa of two or more different males to fertilize the same egg during sexual reproduction. Competition can occur when females have multiple potential mating partners. Greater choice and variety of mates increases a female's chance to produce more viable offspring. However, multiple mates for a female means each individual male has decreased chances of producing offspring. Sperm competition is an evolutionary pressure on males, and has led to the development of adaptations to increase male's chance of reproductive success. Sperm competition results in a sexual conflict between males and females. Males have evolved several defensive tactics including: mate-guarding, mating plugs, and releasing toxic seminal substances to reduce female re-mating tendencies to cope with sperm competition. Offensive tactics of sperm competition involve direct interference by one male on the reproductive success of another male, for instance by physically removing another male's sperm prior to mating with a female. For an example, see Gryllus bimaculatus.

Molecular ecology is a field of evolutionary biology that is concerned with applying molecular population genetics, molecular phylogenetics, and more recently genomics to traditional ecological questions. It is virtually synonymous with the field of "Ecological Genetics" as pioneered by Theodosius Dobzhansky, E. B. Ford, Godfrey M. Hewitt, and others. These fields are united in their attempt to study genetic-based questions "out in the field" as opposed to the laboratory. Molecular ecology is related to the field of conservation genetics.

Animal sexual behaviour takes many different forms, including within the same species. Common mating or reproductively motivated systems include monogamy, polygyny, polyandry, polygamy and promiscuity. Other sexual behaviour may be reproductively motivated or non-reproductively motivated.

Fish reproductive organs include testes and ovaries. In most species, gonads are paired organs of similar size, which can be partially or totally fused. There may also be a range of secondary organs that increase reproductive fitness. The genital papilla is a small, fleshy tube behind the anus in some fishes, from which the sperm or eggs are released; the sex of a fish can often be determined by the shape of its papilla.
Inbreeding depression is the reduced biological fitness that has the potential to result from inbreeding. Biological fitness refers to an organism's ability to survive and perpetuate its genetic material. Inbreeding depression is often the result of a population bottleneck. In general, the higher the genetic variation or gene pool within a breeding population, the less likely it is to suffer from inbreeding depression, though inbreeding and outbreeding depression can simultaneously occur.
Philopatry is the tendency of an organism to stay in or habitually return to a particular area. The causes of philopatry are numerous, but natal philopatry, where animals return to their birthplace to breed, may be the most common. The term derives from the Greek roots philo, "liking, loving" and patra, "fatherland", although in recent years the term has been applied to more than just the animal's birthplace. Recent usage refers to animals returning to the same area to breed despite not being born there, and migratory species that demonstrate site fidelity: reusing stopovers, staging points, and wintering grounds.
Canine reproduction is the process of sexual reproduction in domestic dogs, wolves, coyotes and other canine species.
Cooperative breeding is a social system characterized by alloparental care: offspring receive care not only from their parents, but also from additional group members, often called helpers. Cooperative breeding encompasses a wide variety of group structures, from a breeding pair with helpers that are offspring from a previous season, to groups with multiple breeding males and females (polygynandry) and helpers that are the adult offspring of some but not all of the breeders in the group, to groups in which helpers sometimes achieve co-breeding status by producing their own offspring as part of the group's brood. Cooperative breeding occurs across taxonomic groups including birds, mammals, fish, and insects.

Stegodyphus lineatus is the only European species of the spider genus Stegodyphus. Male S. lineatus can grow up to 12 mm long while females can grow up to 15 mm. The colour can range from whitish to almost black. In most individuals the opisthosoma is whitish with two broad black longitudinal stripes. Males and females look similar, but the male is generally richer in contrast and has a bulbous forehead. The species name refers to the black lines on the back of these spiders. S. lineatus is found in the southern Mediterranean region of Europe and as far east as Tajikistan.
Kin recognition, also called kin detection, is an organism's ability to distinguish between close genetic kin and non-kin. In evolutionary biology and psychology, such an ability is presumed to have evolved for inbreeding avoidance, though animals do not typically avoid inbreeding.
Bateman's principle, in evolutionary biology, is that in most species, variability in reproductive success is greater in males than in females. It was first proposed by Angus John Bateman (1919–1996), an English geneticist. Bateman suggested that, since males are capable of producing millions of sperm cells with little effort, while females invest much higher levels of energy in order to nurture a relatively small number of eggs, the female plays a significantly larger role in their offspring's reproductive success. Bateman's paradigm thus views females as the limiting factor of parental investment, over which males will compete in order to copulate successfully.

Eusociality evolved repeatedly in different orders of animals, notably termites and the Hymenoptera. This 'true sociality' in animals, in which sterile individuals work to further the reproductive success of others, is found in termites, ambrosia beetles, gall-dwelling aphids, thrips, marine sponge-dwelling shrimp, naked mole-rats, and many genera in the insect order Hymenoptera. The fact that eusociality has evolved so often in the Hymenoptera, but remains rare throughout the rest of the animal kingdom, has made its evolution a topic of debate among evolutionary biologists. Eusocial organisms at first appear to behave in stark contrast with simple interpretations of Darwinian evolution: passing on one's genes to the next generation, or fitness, is a central idea in evolutionary biology.

The term seabird is used for many families of birds in several orders that spend the majority of their lives at sea. Seabirds make up some, if not all, of the families in the following orders: Procellariiformes, Sphenisciformes, Pelecaniformes, and Charadriiformes. Many seabirds remain at sea for several consecutive years at a time, without ever seeing land. Breeding is the central purpose for seabirds to visit land. The breeding period is usually extremely protracted in many seabirds and may last over a year in some of the larger albatrosses; this is in stark contrast with passerine birds. Seabirds nest in single or mixed-species colonies of varying densities, mainly on offshore islands devoid of terrestrial predators. However, seabirds exhibit many unusual breeding behaviors during all stages of the reproductive cycle that are not extensively reported outside of the primary scientific literature.

The major histocompatibility complex in sexual selection concerns how major histocompatibility complex (MHC) molecules allow for immune system surveillance of the population of protein molecules in a host's cells. In 1976, Yamazaki et al. demonstrated a sexual selection mate choice by male mice for females of a different MHC.

Sexual selection in insects is about how sexual selection functions in insects. The males of some species have evolved exaggerated adornments and mechanisms for self-defense. These traits play a role in increasing male reproductive expectations by triggering male-male competition or influencing the female mate choice, and can be thought of as functioning on three different levels: individuals, colonies, and populations within an area.
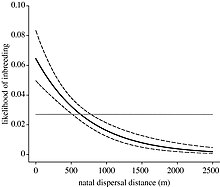
Inbreeding in fish is the mating of closely related individuals, leading to an increase in homozygosity. Repeated inbreeding generally leads to morphological abnormalities and a reduction in fitness in the offspring. In the wild, fish have a number of ways to avoid inbreeding, both before and after copulation.

Primate sociality is an area of primatology that aims to study the interactions between three main elements of a primate social network: the social organisation, the social structure and the mating system. The intersection of these three structures describe the socially complex behaviours and relationships occurring among adult males and females of a particular species. Cohesion and stability of groups are maintained through a confluence of factors, including: kinship, willingness to cooperate, frequency of agonistic behaviour, or varying intensities of dominance structures.