Related Research Articles

Sexual selection is a mode of natural selection in which members of one biological sex choose mates of the other sex to mate with, and compete with members of the same sex for access to members of the opposite sex. These two forms of selection mean that some individuals have greater reproductive success than others within a population, for example because they are more attractive or prefer more attractive partners to produce offspring. Successful males benefit from frequent mating and monopolizing access to one or more fertile females. Females can maximise the return on the energy they invest in reproduction by selecting and mating with the best males.

Sexual dimorphism is the condition where sexes of the same species exhibit different morphological characteristics, particularly characteristics not directly involved in reproduction. The condition occurs in most animals and some plants. Differences may include secondary sex characteristics, size, weight, color, markings, or behavioral or cognitive traits. Male–male reproductive competition has evolved a diverse array of sexually dimorphic traits. Aggressive utility traits such as "battle" teeth and blunt heads reinforced as battering rams are used as weapons in aggressive interactions between rivals. Passive displays such as ornamental feathering or song-calling have also evolved mainly through sexual selection. These differences may be subtle or exaggerated and may be subjected to sexual selection and natural selection. The opposite of dimorphism is monomorphism, when both biological sexes are phenotypically indistinguishable from each other.
Population genetics is a subfield of genetics that deals with genetic differences within and among populations, and is a part of evolutionary biology. Studies in this branch of biology examine such phenomena as adaptation, speciation, and population structure.

Behavioral ecology, also spelled behavioural ecology, is the study of the evolutionary basis for animal behavior due to ecological pressures. Behavioral ecology emerged from ethology after Niko Tinbergen outlined four questions to address when studying animal behaviors: What are the proximate causes, ontogeny, survival value, and phylogeny of a behavior?

Pleiotropy occurs when one gene influences two or more seemingly unrelated phenotypic traits. Such a gene that exhibits multiple phenotypic expression is called a pleiotropic gene. Mutation in a pleiotropic gene may have an effect on several traits simultaneously, due to the gene coding for a product used by a myriad of cells or different targets that have the same signaling function.
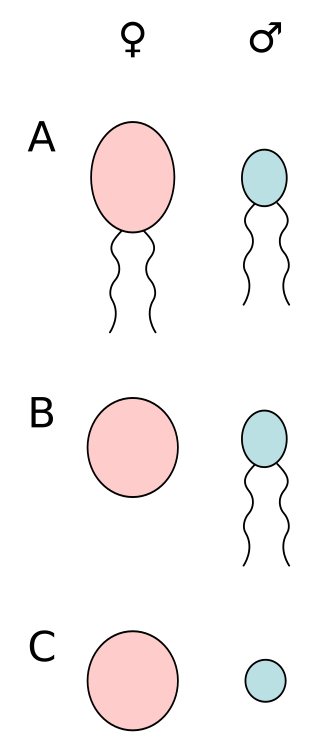
Anisogamy is a form of sexual reproduction that involves the union or fusion of two gametes that differ in size and/or form. The smaller gamete is male, a sperm cell, whereas the larger gamete is female, typically an egg cell. Anisogamy is predominant among multicellular organisms. In both plants and animals gamete size difference is the fundamental difference between females and males.

Molecular ecology is a field of evolutionary biology that is concerned with applying molecular population genetics, molecular phylogenetics, and more recently genomics to traditional ecological questions. It is virtually synonymous with the field of "Ecological Genetics" as pioneered by Theodosius Dobzhansky, E. B. Ford, Godfrey M. Hewitt, and others. These fields are united in their attempt to study genetic-based questions "out in the field" as opposed to the laboratory. Molecular ecology is related to the field of conservation genetics.

Parental investment, in evolutionary biology and evolutionary psychology, is any parental expenditure that benefits offspring. Parental investment may be performed by both males and females, females alone or males alone. Care can be provided at any stage of the offspring's life, from pre-natal to post-natal.
Interlocus contest evolution (ICE) is a process of intergenomic conflict by which different loci within a single genome antagonistically coevolve. ICE supposes that the Red Queen process, which is characterized by a never-ending antagonistic evolutionary arms race, does not only apply to species but also to genes within the genome of a species.
Monogamous pairing in animals refers to the natural history of mating systems in which species pair bond to raise offspring. This is associated, usually implicitly, with sexual monogamy.

Sexual conflict or sexual antagonism occurs when the two sexes have conflicting optimal fitness strategies concerning reproduction, particularly over the mode and frequency of mating, potentially leading to an evolutionary arms race between males and females. In one example, males may benefit from multiple matings, while multiple matings may harm or endanger females, due to the anatomical differences of that species. Sexual conflict underlies the evolutionary distinction between male and female.

Mate choice is one of the primary mechanisms under which evolution can occur. It is characterized by a "selective response by animals to particular stimuli" which can be observed as behavior. In other words, before an animal engages with a potential mate, they first evaluate various aspects of that mate which are indicative of quality—such as the resources or phenotypes they have—and evaluate whether or not those particular trait(s) are somehow beneficial to them. The evaluation will then incur a response of some sort.

The sexy son hypothesis in evolutionary biology and sexual selection, proposed by Patrick J. Weatherhead and Raleigh J. Robertson of Queen's University in Kingston, Ontario in 1979, states that a female's ideal mate choice among potential mates is one whose genes will produce males with the best chance of reproductive success. This implies that other benefits the father can offer the mother or offspring are less relevant than they may appear, including his capacity as a parental caregiver, territory and any nuptial gifts. Fisher's principle means that the sex ratio is always near 1:1 between males and females, yet what matters most are her "sexy sons'" future breeding successes, more likely if they have a promiscuous father, in creating large numbers of offspring carrying copies of her genes. This sexual selection hypothesis has been researched in species such as the European pied flycatcher.
Sex-limited genes are genes that are present in both sexes of sexually reproducing species but are expressed in only one sex and have no penetrance, or are simply 'turned off' in the other. In other words, sex-limited genes cause the two sexes to show different traits or phenotypes, despite having the same genotype. This term is restricted to autosomal traits, and should not be confused with sex-linked characteristics, which have to do with genetic differences on the sex chromosomes. Sex-limited genes are also distinguished from sex-influenced genes, where the same gene will show differential expression in each sex. Sex-influenced genes commonly show a dominant/recessive relationship, where the same gene will have a dominant effect in one sex and a recessive effect in the other. However, the resulting phenotypes caused by sex-limited genes are present in only one sex and can be seen prominently in various species that typically show high sexual dimorphism.
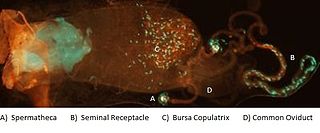
Female sperm storage is a biological process and often a type of sexual selection in which sperm cells transferred to a female during mating are temporarily retained within a specific part of the reproductive tract before the oocyte, or egg, is fertilized. This process takes place in some species of animals, but not in humans. The site of storage is variable among different animal taxa and ranges from structures that appear to function solely for sperm retention, such as insect spermatheca and bird sperm storage tubules, to more general regions of the reproductive tract enriched with receptors to which sperm associate before fertilization, such as the caudal portion of the cow oviduct containing sperm-associating annexins. Female sperm storage is an integral stage in the reproductive process for many animals with internal fertilization. It has several documented biological functions including:
Sexual antagonistic co-evolution is the relationship between males and females where sexual morphology changes over time to counteract the opposite's sex traits to achieve the maximum reproductive success. This has been compared to an arms race between sexes. In many cases, male mating behavior is detrimental to the female's fitness. For example, when insects reproduce by means of traumatic insemination, it is very disadvantageous to the female's health. During mating, males will try to inseminate as many females as possible, however, the more times a female's abdomen is punctured, the less likely she is to survive. Females that possess traits to avoid multiple matings will be more likely to survive, resulting in a change in morphology. In males, genitalia is relatively simple and more likely to vary among generations compared to female genitalia. This results in a new trait that females have to avoid in order to survive.
Interlocus sexual conflict is a type of sexual conflict that occurs through the interaction of a set of antagonistic alleles at two or more different loci, or the location of a gene on a chromosome, in males and females, resulting in the deviation of either or both sexes from the fitness optima for the traits. A co-evolutionary arms race is established between the sexes in which either sex evolves a set of antagonistic adaptations that is detrimental to the fitness of the other sex. The potential for reproductive success in one organism is strengthened while the fitness of the opposite sex is weakened. Interlocus sexual conflict can arise due to aspects of male–female interactions such as mating frequency, fertilization, relative parental effort, female remating behavior, and female reproductive rate.

Sexual selection in scaled reptiles studies how sexual selection manifests in snakes and lizards, which constitute the order Squamata of reptiles. Each of the over three thousand snakes use different tactics in acquiring mates. Ritual combat between males for the females they want to mate with includes topping, a behavior exhibited by most viperids in which one male will twist around the vertically elevated fore body of its opponent and forcing it downward. It is common for neck biting to occur while the snakes are entwined.

Reinforcement is a process of speciation where natural selection increases the reproductive isolation between two populations of species. This occurs as a result of selection acting against the production of hybrid individuals of low fitness. The idea was originally developed by Alfred Russel Wallace and is sometimes referred to as the Wallace effect. The modern concept of reinforcement originates from Theodosius Dobzhansky. He envisioned a species separated allopatrically, where during secondary contact the two populations mate, producing hybrids with lower fitness. Natural selection results from the hybrid's inability to produce viable offspring; thus members of one species who do not mate with members of the other have greater reproductive success. This favors the evolution of greater prezygotic isolation. Reinforcement is one of the few cases in which selection can favor an increase in prezygotic isolation, influencing the process of speciation directly. This aspect has been particularly appealing among evolutionary biologists.
Genetic incompatibility describes the process by which mating yields offspring that are nonviable, prone to disease, or genetically defective in some way. In nature, animals can ill afford to devote costly resources for little or no reward, ergo, mating strategies have evolved to allow females to choose or otherwise determine mates which are more likely to result in viable offspring.
References
- ↑ Pennell, Tanya; Morrow, Edward H (2013). "Two sexes, one genome: the evolutionary dynamics of intralocus sexual conflict". Ecology and Evolution. 3 (6): 1819–34. doi: 10.1002/ece3.540 . PMC 3686212 . PMID 23789088.
- ↑ Chapman, T; Arnqvist, G; Bangham, J; Rowe, L (2003). "Sexual conflict". Trends in Ecology and Evolution. 18: 41–47. doi:10.1016/s0169-5347(02)00004-6.
- ↑ Parker, G.A. (1979), "Sexual Selection and Sexual Conflict", Sexual Selection and Reproductive Competition in Insects, Elsevier, pp. 123–166, doi:10.1016/b978-0-12-108750-0.50010-0, ISBN 9780121087500
- ↑ Bonduriansky, Russell; Chenoweth, Stephen F (2009). "Intralocus sexual conflict". Trends in Ecology and Evolution. 24 (5): 280–8. doi:10.1016/j.tree.2008.12.005. PMID 19307043.
- ↑ Connallon, Tim; Hall, Matthew D (2018). "Genetic constraints on adaptation: a theoretical primer for the genomics era". Annals of the New York Academy of Sciences. 1422 (1): 65–87. Bibcode:2018NYASA1422...65C. doi: 10.1111/nyas.13536 . PMID 29363779.
- ↑ Matthews, Genevieve; Hangartner, Sandra; Chapple, David G; Connallon, Tim (2019). "Quantifying maladaptation during the evolution of sexual dimorphism". Proceedings of the Royal Society B: Biological Sciences. 286 (1908): 20191372. doi: 10.1098/rspb.2019.1372 . PMC 6710593 . PMID 31409252.
- ↑ Rice, William R (1984). "Sex Chromosomes and the Evolution of Sexual Dimorphism". Evolution. 38 (4): 735–742. doi: 10.1111/j.1558-5646.1984.tb00346.x . PMID 28555827.
- ↑ Guerrero, Rafael F; Kirkpatrick, Mark (2014). "Signatures of sex-antagonistic selection on recombining sex chromosomes". Genetics. 197 (2): 531–41. doi: 10.1534/genetics.113.156026 . PMC 4063913 . PMID 24578352.
- ↑ Connallon, Tim; Clark, Andrew (2012). "A general population genetic framework for antagonistic selection that accounts for demography and recurrent mutation". Genetics. 190 (4): 1477–89. doi: 10.1534/genetics.111.137117 . PMC 3316657 . PMID 22298707.
- 1 2 Doorn, Van; Sander, G. (2009). "Intralocus Sexual Conflict". Annals of the New York Academy of Sciences. 1168 (1): 52–71. Bibcode:2009NYASA1168...52V. doi:10.1111/j.1749-6632.2009.04573.x. PMID 19566703.
- ↑ Mills, S. C.; Koskela, E.; Mappes, T. (2011). "Intralocus Sexual Conflict for Fitness: Sexually Antagonistic Alleles for Testosterone". Proceedings of the Royal Society B: Biological Sciences. 279 (1735): 1889–95. doi:10.1098/rspb.2011.2340. PMC 3311893 . PMID 22171083.
- ↑ Bielak; Plesnar, Agata; Skrzynecka, Anna M.; Miler, Krzysztof; Radwan, Jacek (2014). "Selection for Alternative Male Reproductive Tactics Alters Intralocus Sexual Conflict". Evolution. 68 (7): 2137–44. doi:10.1111/evo.12409. PMID 24641007. S2CID 30131544.
- ↑ Bonduriansky, R; Chenowith, SF (2009). "Intralocus sexual conflict". Trends in Ecology and Evolution. 24 (5): 280–288. doi:10.1016/j.tree.2008.12.005. PMID 19307043.
- 1 2 Stulp, G.; et al. (2012). "Intralocus Sexual Conflict over Human Height". Biology Letters. 8 (6): 976–78. doi:10.1098/rsbl.2012.0590. PMC 3497124 . PMID 22875819.
- ↑ Pischedda, Alison; Chippindale, Adam K. (2006). "Intralocus Sexual Conflict Diminishes the Benefits of Sexual Selection". PLOS Biology. 4 (11): 2099–2103. doi:10.1371/journal.pbio.0040356. PMC 1618422 . PMID 17105343.
- 1 2 Garver-Apgar, Christine E.; Melissa; Eaton, Joshua M. Tybur; Emery Thompson, Melissa (2011). "Evidence of Intralocus Sexual Conflict: Physically and Hormonally Masculine Individuals Have More Attractive Brothers Relative to Sisters". Evolution and Human Behavior. 32 (6): 423–32. doi:10.1016/j.evolhumbehav.2011.03.005.
- ↑ Rice, WR; Chippindale, AK (2001). "Intersexual ontogenetic conflict". J. Evol. Biol. 14 (5): 685–693. doi: 10.1046/j.1420-9101.2001.00319.x . S2CID 83464823.
- ↑ Flanagan, Sarah P.; Jones, Adam G (2017). "Genome-wide selection components analysis in a fish with male pregnancy". Evolution. 71 (4): 1096–1105. doi:10.1111/evo.13173. PMID 28067418.
- 1 2 Vaux, Felix; Rasmuson, Leif K.; Kautzi, Lisa A.; Rankin, Polly S.; Blume, Matthew T.O.; Lawrence, Kelly A.; Bohn, Sandra; O'Malley, Kathleen G. (2019). "Sex matters: Otolith shape and genomic variation in deacon rockfish (Sebastes diaconus)". Ecology and Evolution. 9 (23): 13153–13173. doi: 10.1002/ece3.5763 . PMC 6912905 . PMID 31871636.
- 1 2 3 Pennell, Tanya M.; Morrow, Edward H. (2013). "Two Sexes, One Genome: The Evolutionary Dynamics of Intralocus Sexual Conflict". Ecology and Evolution. 3 (6): 1819–34. doi:10.1002/ece3.540. PMC 3686212 . PMID 23789088.