
In ecology, a niche is the match of a species to a specific environmental condition. It describes how an organism or population responds to the distribution of resources and competitors and how it in turn alters those same factors. "The type and number of variables comprising the dimensions of an environmental niche vary from one species to another [and] the relative importance of particular environmental variables for a species may vary according to the geographic and biotic contexts".

In ecology, edge effects are changes in population or community structures that occur at the boundary of two or more habitats. Areas with small habitat fragments exhibit especially pronounced edge effects that may extend throughout the range. As the edge effects increase, the boundary habitat allows for greater biodiversity.
Phylogeography is the study of the historical processes that may be responsible for the past to present geographic distributions of genealogical lineages. This is accomplished by considering the geographic distribution of individuals in light of genetics, particularly population genetics.
Allopatric speciation – also referred to as geographic speciation, vicariant speciation, or its earlier name the dumbbell model – is a mode of speciation that occurs when biological populations become geographically isolated from each other to an extent that prevents or interferes with gene flow.

An environmental gradient, or climate gradient, is a change in abiotic (non-living) factors through space. Environmental gradients can be related to factors such as altitude, depth, temperature, soil humidity and precipitation. Often times, a multitude of biotic (living) factors are closely related to these gradients; as a result of a change in an environmental gradient, factors such as species abundance, population density, morphology, primary productivity, predation, and local adaptation may be impacted.
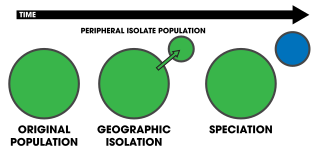
Peripatric speciation is a mode of speciation in which a new species is formed from an isolated peripheral population. Since peripatric speciation resembles allopatric speciation, in that populations are isolated and prevented from exchanging genes, it can often be difficult to distinguish between them. Nevertheless, the primary characteristic of peripatric speciation proposes that one of the populations is much smaller than the other. The terms peripatric and peripatry are often used in biogeography, referring to organisms whose ranges are closely adjacent but do not overlap, being separated where these organisms do not occur—for example on an oceanic island compared to the mainland. Such organisms are usually closely related ; their distribution being the result of peripatric speciation.

Biological dispersal refers to both the movement of individuals from their birth site to their breeding site, as well as the movement from one breeding site to another . Dispersal is also used to describe the movement of propagules such as seeds and spores. Technically, dispersal is defined as any movement that has the potential to lead to gene flow. The act of dispersal involves three phases: departure, transfer, settlement and there are different fitness costs and benefits associated with each of these phases. Through simply moving from one habitat patch to another, the dispersal of an individual has consequences not only for individual fitness, but also for population dynamics, population genetics, and species distribution. Understanding dispersal and the consequences both for evolutionary strategies at a species level, and for processes at an ecosystem level, requires understanding on the type of dispersal, the dispersal range of a given species, and the dispersal mechanisms involved. Biological dispersal can be correlated to population density. The range of variations of a species' location determines expansion range.

Habitat fragmentation describes the emergence of discontinuities (fragmentation) in an organism's preferred environment (habitat), causing population fragmentation and ecosystem decay. Causes of habitat fragmentation include geological processes that slowly alter the layout of the physical environment, and human activity such as land conversion, which can alter the environment much faster and causes the extinction of many species. More specifically, habitat fragmentation is a process by which large and contiguous habitats get divided into smaller, isolated patches of habitats.

The chalkhill blue is a butterfly in the family Lycaenidae. It is a small butterfly that can be found throughout the Palearctic realm, where it occurs primarily in grasslands rich in chalk. Males have a pale blue colour, while females are brown. Both have chequered fringes around their wings.

In parapatric speciation, two subpopulations of a species evolve reproductive isolation from one another while continuing to exchange genes. This mode of speciation has three distinguishing characteristics: 1) mating occurs non-randomly, 2) gene flow occurs unequally, and 3) populations exist in either continuous or discontinuous geographic ranges. This distribution pattern may be the result of unequal dispersal, incomplete geographical barriers, or divergent expressions of behavior, among other things. Parapatric speciation predicts that hybrid zones will often exist at the junction between the two populations.

Species richness, or biodiversity, increases from the poles to the tropics for a wide variety of terrestrial and marine organisms, often referred to as the latitudinal diversity gradient. The latitudinal diversity gradient is one of the most widely recognized patterns in ecology. It has been observed to varying degrees in Earth's past. A parallel trend has been found with elevation, though this is less well-studied.
Species distribution, or speciesdispersion, is the manner in which a biological taxon is spatially arranged. The geographic limits of a particular taxon's distribution is its range, often represented as shaded areas on a map. Patterns of distribution change depending on the scale at which they are viewed, from the arrangement of individuals within a small family unit, to patterns within a population, or the distribution of the entire species as a whole (range). Species distribution is not to be confused with dispersal, which is the movement of individuals away from their region of origin or from a population center of high density.

The dainty damselfly, also known as the dainty bluet, is a blue damselfly of the family Coenagrionidae. This is a scarce species found mainly in southern Europe, northern Africa, southwest Asia, and Central America. C. scitulum are Odonata predators that can reach a length of 30–33 mm at maturity and have hind-wing lengths of 15–20 mm. The males and females do exhibit differing features through their colouration making them easily distinguishable. As shown in the photo to the left, segment eight is blue in colour followed by black markings on segment nine, whereas the females are mostly black near the rear with smaller blue markings. Although they are relatively simple to sex, they are easily confused with the common blue damselfly.
A genetic isolate is a population of organisms that has little to no genetic mixing with other organisms of the same species due to geographic isolation or other factors that prevent reproduction. Genetic isolates form new species through an evolutionary process known as speciation. All modern species diversity is a product of genetic isolates and evolution.
Ecological forecasting uses knowledge of physics, ecology and physiology to predict how ecological populations, communities, or ecosystems will change in the future in response to environmental factors such as climate change. The goal of the approach is to provide natural resource managers with information to anticipate and respond to short and long-term climate conditions.

Forest migration is the movement of large seed plant dominated communities in geographical space over time.

Ecological fitting is "the process whereby organisms colonize and persist in novel environments, use novel resources or form novel associations with other species as a result of the suites of traits that they carry at the time they encounter the novel condition". It can be understood as a situation in which a species' interactions with its biotic and abiotic environment seem to indicate a history of coevolution, when in actuality the relevant traits evolved in response to a different set of biotic and abiotic conditions.

Ecological speciation is a form of speciation arising from reproductive isolation that occurs due to an ecological factor that reduces or eliminates gene flow between two populations of a species. Ecological factors can include changes in the environmental conditions in which a species experiences, such as behavioral changes involving predation, predator avoidance, pollinator attraction, and foraging; as well as changes in mate choice due to sexual selection or communication systems. Ecologically-driven reproductive isolation under divergent natural selection leads to the formation of new species. This has been documented in many cases in nature and has been a major focus of research on speciation for the past few decades.

Climate change and invasive species refers to the process of the environmental destabilization caused by climate change. This environmental change facilitates the spread of invasive species — species that are not historically found in a certain region, and often bring about a negative impact to that region's native species. This complex relationship is notable because climate change and invasive species are also considered by the USDA to be two of the top four causes of global biodiversity loss.
In biology, parallel speciation is a type of speciation where there is repeated evolution of reproductively isolating traits via the same mechanisms occurring between separate yet closely related species inhabiting different environments. This leads to a circumstance where independently evolved lineages have developed reproductive isolation from their ancestral lineage, but not from other independent lineages that inhabit similar environments. In order for parallel speciation to be confirmed, there is a set of three requirements that has been established that must be met: there must be phylogenetic independence between the separate populations inhabiting similar environments to ensure that the traits responsible for reproductive isolation evolved separately, there must be reproductive isolation not only between the ancestral population and the descendent population, but also between descendent populations that inhabit dissimilar environments, and descendent populations that inhabit similar environments must not be reproductively isolated from one another. To determine if natural selection specifically is the cause of parallel speciation, a fourth requirement has been established that includes identifying and testing an adaptive mechanism, which eliminates the possibility of a genetic factor such as polyploidy being the responsible agent.


