Related Research Articles

Photosynthesis is a process used by plants and other organisms to convert light energy into chemical energy that can later be released to fuel the organisms' activities. This chemical energy is stored in carbohydrate molecules, such as sugars, which are synthesized from carbon dioxide and water – hence the name photosynthesis, from the Greek phōs (φῶς), "light", and sunthesis (σύνθεσις), "putting together". In most cases, oxygen is also released as a waste product. Most plants, most algae, and cyanobacteria perform photosynthesis; such organisms are called photoautotrophs. Photosynthesis is largely responsible for producing and maintaining the oxygen content of the Earth's atmosphere, and supplies most of the energy necessary for life on Earth.
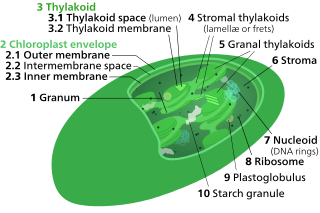
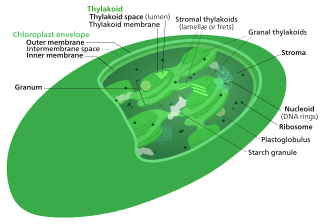
Thylakoids are membrane-bound compartments inside chloroplasts and cyanobacteria. They are the site of the light-dependent reactions of photosynthesis. Thylakoids consist of a thylakoid membrane surrounding a thylakoid lumen. Chloroplast thylakoids frequently form stacks of disks referred to as grana. Grana are connected by intergranal/ stroma thylakoids, which join granum stacks together as a single functional compartment.

Ribulose-1,5-bisphosphate carboxylase-oxygenase, commonly known by the abbreviations RuBisCo, rubisco, RuBPCase, or RuBPco, is an enzyme involved in the first major step of carbon fixation, a process by which the atmospheric carbon dioxide is converted by plants and other photosynthetic organisms to energy-rich molecules such as glucose. In chemical terms, it catalyzes the carboxylation of ribulose-1,5-bisphosphate. It is probably the most abundant enzyme on Earth.
C4 carbon fixation or the Hatch–Slack pathway is one of three known photosynthetic processes of carbon fixation in plants. It owes the names to the discovery by Marshall Davidson Hatch and Charles Roger Slack that some plants, when supplied with 14CO
2, incorporate the 14C label into four-carbon molecules first.

Photorespiration (also known as the oxidative photosynthetic carbon cycle, or C2 photosynthesis) refers to a process in plant metabolism where the enzyme RuBisCO oxygenates RuBP, wasting some of the energy produced by photosynthesis. The desired reaction is the addition of carbon dioxide to RuBP (carboxylation), a key step in the Calvin–Benson cycle, but approximately 25% of reactions by RuBisCO instead add oxygen to RuBP (oxygenation), creating a product that cannot be used within the Calvin–Benson cycle. This process reduces the efficiency of photosynthesis, potentially reducing photosynthetic output by 25% in C3 plants. Photorespiration involves a complex network of enzyme reactions that exchange metabolites between chloroplasts, leaf peroxisomes and mitochondria.

Reactive oxygen species (ROS) are highly reactive chemical molecules formed due to the electron acceptability of O2. Examples of ROS include peroxides, superoxide, hydroxyl radical, singlet oxygen, and alpha-oxygen.
The Calvin cycle,light-independent reactions, bio synthetic phase,dark reactions, or photosynthetic carbon reduction (PCR) cycle of photosynthesis are the chemical reactions that convert carbon dioxide and other compounds into glucose. These reactions occur in the stroma, the fluid-filled area of a chloroplast outside the thylakoid membranes. These reactions take the products of light-dependent reactions and perform further chemical processes on them. The Calvin cycle uses the reducing powers ATP and NADPH from the light dependent reactions to produce sugars for the plant to use. These substrates are used in a series of reduction-oxidation reactions to produce sugars in a step-wise process. There is no direct reaction that converts CO2 to a sugar because all of the energy would be lost to heat.] There are three phases to the light-independent reactions, collectively called the Calvin cycle: carbon fixation, reduction reactions, and ribulose 1,5-bisphosphate (RuBP) regeneration.

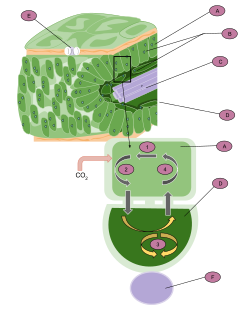
Photosystems are functional and structural units of protein complexes involved in photosynthesis that together carry out the primary photochemistry of photosynthesis: the absorption of light and the transfer of energy and electrons. Photosystems are found in the thylakoid membranes of plants, algae and cyanobacteria which are located inside the chloroplasts of plants and algae, and in the cytoplasmic membrane of photosynthetic bacteria. There are two kinds of photosystems: I and II.
Ferredoxins are iron–sulfur proteins that mediate electron transfer in a range of metabolic reactions. The term "ferredoxin" was coined by D.C. Wharton of the DuPont Co. and applied to the "iron protein" first purified in 1962 by Mortenson, Valentine, and Carnahan from the anaerobic bacterium Clostridium pasteurianum.

The cytochrome b6f complex is an enzyme found in the thylakoid membrane in chloroplasts of plants, cyanobacteria, and green algae, that catalyzes the transfer of electrons from plastoquinol to plastocyanin. The reaction is analogous to the reaction catalyzed by cytochrome bc1 of the mitochondrial electron transport chain. During photosynthesis, the cytochrome b6f complex is one step along the chain that transfers electrons from Photosystem II to Photosystem I, and at the same time pumps protons into the thylakoid space that contribute to create an electrochemical (energy) gradient which is later used to synthesize ATP from ADP.

A photosynthetic reaction center is a complex of several proteins, pigments and other co-factors that together execute the primary energy conversion reactions of photosynthesis. Molecular excitations, either originating directly from sunlight or transferred as excitation energy via light-harvesting antenna systems, give rise to electron transfer reactions along the path of a series of protein-bound co-factors. These co-factors are light-absorbing molecules such as chlorophyll and phaeophytin, as well as quinones. The energy of the photon is used to excite an electron of a pigment. The free energy created is then used to reduce a chain of nearby electron acceptors, which have progressively higher redox-potentials. These electron transfer steps are the initial phase of a series of energy conversion reactions, ultimately resulting in the conversion of the energy of photons to the storage of that energy by the production of chemical bonds.

DCMU is an algicide and herbicide of the arylurea class that inhibits photosynthesis. It was introduced by Bayer in 1954 under the trade name of Diuron.
In enzymology, a ferredoxin-NADP+ reductase (EC 1.18.1.2) abbreviated FNR, is an enzyme that catalyzes the chemical reaction

Sedoheptulose-bisphosphatase is an enzyme that catalyzes the removal of a phosphate group from sedoheptulose 1,7-bisphosphate to produce sedoheptulose 7-phosphate. SBPase is an example of a phosphatase, or, more generally, a hydrolase. This enzyme participates in the Calvin cycle.
Dioxygen plays an important role in the energy metabolism of living organisms. Free oxygen is produced in the biosphere through photolysis of water during photosynthesis in cyanobacteria, green algae, and plants. During oxidative phosphorylation in cellular respiration, the chemical energy of oxygen is released as it is reduced to water, thus closing the biological water-oxygen redox cycle.
Foyer-Halliwell-Asada pathway
In photosynthesis, the light-dependent reactions take place on the thylakoid membranes. The inside of the thylakoid membrane is called the lumen, and outside the thylakoid membrane is the stroma, where the light-independent reactions take place. The thylakoid membrane contains some integral membrane protein complexes that catalyze the light reactions. There are four major protein complexes in the thylakoid membrane: Photosystem II (PSII), Cytochrome b6f complex, Photosystem I (PSI), and ATP synthase. These four complexes work together to ultimately create the products ATP and NADPH.
All living cells produce reactive oxygen species (ROS) as a byproduct of metabolism. ROS are reduced oxygen intermediates that include the superoxide radical (O2−) and the hydroxyl radical (OH•), as well as the non-radical species hydrogen peroxide (H2O2). These ROS are important in the normal functioning of cells, playing a role in signal transduction and the expression of transcription factors. However, when present in excess, ROS can cause damage to proteins, lipids and DNA by reacting with these biomolecules to modify or destroy their intended function. As an example, the occurrence of ROS have been linked to the aging process in humans, as well as several other diseases including Alzheimer's, rheumatoid arthritis, Parkinson's, and some cancers. Their potential for damage also makes reactive oxygen species useful in direct protection from invading pathogens, as a defense response to physical injury, and as a mechanism for stopping the spread of bacteria and viruses by inducing programmed cell death.
Plastid terminal oxidase or plastoquinol terminal oxidase (PTOX) is an enzyme that resides on the thylakoid membranes of plant and algae chloroplasts and on the membranes of cyanobacteria. The enzyme was hypothesized to exist as a photosynthetic oxidase in 1982 and was verified by sequence similarity to the mitochondrial alternative oxidase (AOX). The two oxidases evolved from a common ancestral protein in prokaryotes, and they are so functionally and structurally similar that a thylakoid-localized AOX can restore the function of a PTOX knockout.
The Hill reaction is the light-driven transfer of electrons from water to Hill reagents in a direction against the chemical potential gradient as part of photosynthesis. Robin Hill discovered the reaction in 1937. He demonstrated that the process by which plants produce oxygen is separate from the process that converts carbon dioxide to sugars.
References
- ↑ Mehler, Alan (1951). "Studies on reactions of illuminated chloroplasts: I. Mechanism of the reduction of oxygen and other hill reagents". Archives of Biochemistry and Biophysics. 33 (1): 65–77. doi:10.1016/0003-9861(51)90082-3.
- ↑ Asada, Kozi (June 1999). "The Water-Cycle in Chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons". Annual Review of Plant Physiology and Plant Molecular Biology. 50: 601-639. doi:10.1146/annurev.arplant.50.1.601.
- ↑ Mano, Endo and Miyake (2016). "How do photosynthetic organisms manage light stress? A tribute to the late Professor Kozi Asada". Plant and Cell Physiology. 57 (7): 1351-1353. doi: 10.1093/pcp/pcw116 .
- ↑ Badger, M.; von Caemmerer, S.; Ruuska, S.; Nakano, H. (2000). "Electron flow to oxygen in higher plants and algae: rates and control of direct photoreduction (Mehler reaction) and rubisco oxygenase". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 355 (1402): 1433–1446. doi:10.1098/rstb.2000.0704. PMC 1692866 . PMID 11127997.
- ↑ Ruuska, S.A.; Badger, M.R.; von Caemmerer, S. (2000). "Photosynthetic electron sinks in transgenic tobacco with reduced amounts of Rubisco: little evidence for significant Mehler reaction". Journal of Experimental Botany. 51: 357–368. doi: 10.1093/jexbot/51.suppl_1.357 .
- ↑ Driever, S.M.; Baker, N. (2011). "The water–water cycle in leaves is not a major alternative electron sink for dissipation of excess excitation energy when CO
2 assimilation is restricted". Plant, Cell & Environment. 34 (5): 837–846. doi:10.1111/j.1365-3040.2011.02288.x. - ↑ Heber, Ulrich (2002-01-01). "Irrungen, Wirrungen? The Mehler reaction in relation to cyclic electron transport in C3 plants". Photosynthesis Research. 73 (1–3): 223–231. doi:10.1023/A:1020459416987. ISSN 1573-5079. PMID 16245125.