
Molybdenum is an essential element in most organisms. [1] It is most notably present in nitrogenase [2] which is an essential part of nitrogen fixation. [3] [4]
Molybdenum is an essential element in most organisms. [1] It is most notably present in nitrogenase [2] which is an essential part of nitrogen fixation. [3] [4]
Molybdenum is an essential element in most organisms; a 2008 research paper speculated that a scarcity of molybdenum in the Earth's early oceans may have strongly influenced the evolution of eukaryotic life (which includes all plants and animals). [1]
At least 50 molybdenum-containing enzymes have been identified, mostly in bacteria. [5] [6] Those enzymes include aldehyde oxidase, sulfite oxidase and xanthine oxidase. [7] With one exception, Mo in proteins is bound by molybdopterin to give the molybdenum cofactor. The only known exception is nitrogenase, which uses the FeMoco cofactor, which has the formula Fe7MoS9C. [8]
In terms of function, molybdoenzymes catalyze the oxidation and sometimes reduction of certain small molecules in the process of regulating nitrogen, sulfur, and carbon. [9] In some animals, and in humans, the oxidation of xanthine to uric acid, a process of purine catabolism, is catalyzed by xanthine oxidase, a molybdenum-containing enzyme. The activity of xanthine oxidase is directly proportional to the amount of molybdenum in the body. An extremely high concentration of molybdenum reverses the trend and can inhibit purine catabolism and other processes. Molybdenum concentration also affects protein synthesis, metabolism, and growth. [10]
Mo is a component in most nitrogenases. Among molybdoenzymes, nitrogenases are unique in lacking the molybdopterin. [11] [12] Nitrogenases catalyze the production of ammonia from atmospheric nitrogen:
The biosynthesis of the FeMoco active site is highly complex. [13]


Molybdate is transported in the body as MoO42−. [10]
Molybdenum is an essential trace dietary element. [14] Four mammalian Mo-dependent enzymes are known, all of them harboring a pterin-based molybdenum cofactor (Moco) in their active site: sulfite oxidase, xanthine oxidoreductase, aldehyde oxidase, and mitochondrial mitochondrial amidoxime reductase. [15] People severely deficient in molybdenum have poorly functioning sulfite oxidase and are prone to toxic reactions to sulfites in foods. [16] [17] The human body contains about 0.07 mg of molybdenum per kilogram of body weight, [18] with higher concentrations in the liver and kidneys and lower in the vertebrae. [19] Molybdenum is also present within human tooth enamel and may help prevent its decay. [20]
Acute toxicity has not been seen in humans, and the toxicity depends strongly on the chemical state. Studies on rats show a median lethal dose (LD50) as low as 180 mg/kg for some Mo compounds. [21] Although human toxicity data is unavailable, animal studies have shown that chronic ingestion of more than 10 mg/day of molybdenum can cause diarrhea, growth retardation, infertility, low birth weight, and gout; it can also affect the lungs, kidneys, and liver. [22] [23] Sodium tungstate is a competitive inhibitor of molybdenum. Dietary tungsten reduces the concentration of molybdenum in tissues. [19]
Low soil concentration of molybdenum in a geographical band from northern China to Iran results in a general dietary molybdenum deficiency, and is associated with increased rates of esophageal cancer. [24] [25] [26] Compared to the United States, which has a greater supply of molybdenum in the soil, people living in those areas have about 16 times greater risk for esophageal squamous cell carcinoma. [27]
Molybdenum deficiency has also been reported as a consequence of non-molybdenum supplemented total parenteral nutrition (complete intravenous feeding) for long periods of time. It results in high blood levels of sulfite and urate, in much the same way as molybdenum cofactor deficiency. Since pure molybdenum deficiency from this cause occurs primarily in adults, the neurological consequences are not as marked as in cases of congenital cofactor deficiency. [28]
A congenital molybdenum cofactor deficiency disease, seen in infants, is an inability to synthesize molybdenum cofactor, the heterocyclic molecule discussed above that binds molybdenum at the active site in all known human enzymes that use molybdenum. The resulting deficiency results in high levels of sulfite and urate, and neurological damage. [29] [30]
Most molybdenum is excreted from the human body as molybdate in the urine. Furthermore, urinary excretion of molybdenum increases as dietary molybdenum intake increases. Small amounts of molybdenum are excreted from the body in the feces by way of the bile; small amounts also can be lost in sweat and in hair. [31] [32]
High levels of molybdenum can interfere with the body's uptake of copper, producing copper deficiency. Molybdenum prevents plasma proteins from binding to copper, and it also increases the amount of copper that is excreted in urine. Ruminants that consume high levels of molybdenum suffer from diarrhea, stunted growth, anemia, and achromotrichia (loss of fur pigment). These symptoms can be alleviated by copper supplements, either dietary or injection. [33] The effective copper deficiency can be aggravated by excess sulfur. [19] [34]
Copper reduction or deficiency can also be deliberately induced for therapeutic purposes by the compound ammonium tetrathiomolybdate, in which the bright red anion tetrathiomolybdate is the copper-chelating agent. Tetrathiomolybdate was first used therapeutically in the treatment of copper toxicosis in animals. It was then introduced as a treatment in Wilson's disease, a hereditary copper metabolism disorder in humans; it acts both by competing with copper absorption in the bowel and by increasing excretion. It has also been found to have an inhibitory effect on angiogenesis, potentially by inhibiting the membrane translocation process that is dependent on copper ions. [35] This is a promising avenue for investigation of treatments for cancer, age-related macular degeneration, and other diseases that involve a pathologic proliferation of blood vessels. [36] [37]
In some grazing livestock, most strongly in cattle, molybdenum excess in the soil of pasturage can produce scours (diarrhea) if the pH of the soil is neutral to alkaline; see teartness.

Molybdenum is a chemical element; it has symbol Mo and atomic number 42. The name derived from Ancient Greek Μόλυβδος molybdos, meaning lead, since its ores were confused with lead ores. Molybdenum minerals have been known throughout history, but the element was discovered in 1778 by Carl Wilhelm Scheele. The metal was first isolated in 1781 by Peter Jacob Hjelm.

Uric acid is a heterocyclic compound of carbon, nitrogen, oxygen, and hydrogen with the formula C5H4N4O3. It forms ions and salts known as urates and acid urates, such as ammonium acid urate. Uric acid is a product of the metabolic breakdown of purine nucleotides, and it is a normal component of urine. High blood concentrations of uric acid can lead to gout and are associated with other medical conditions, including diabetes and the formation of ammonium acid urate kidney stones.
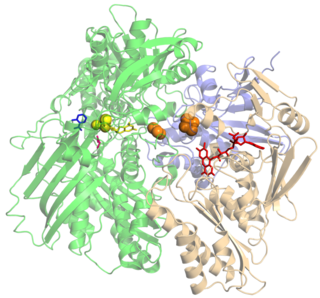
Xanthine oxidase is a form of xanthine oxidoreductase, a type of enzyme that generates reactive oxygen species. These enzymes catalyze the oxidation of hypoxanthine to xanthine and can further catalyze the oxidation of xanthine to uric acid. These enzymes play an important role in the catabolism of purines in some species, including humans.

A catecholamine is a monoamine neurotransmitter, an organic compound that has a catechol and a side-chain amine.

Metalloprotein is a generic term for a protein that contains a metal ion cofactor. A large proportion of all proteins are part of this category. For instance, at least 1000 human proteins contain zinc-binding protein domains although there may be up to 3000 human zinc metalloproteins.
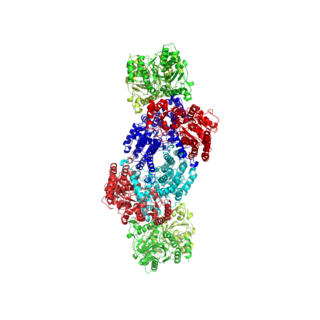
Nitrogenases are enzymes (EC 1.18.6.1EC 1.19.6.1) that are produced by certain bacteria, such as cyanobacteria (blue-green bacteria) and rhizobacteria. These enzymes are responsible for the reduction of nitrogen (N2) to ammonia (NH3). Nitrogenases are the only family of enzymes known to catalyze this reaction, which is a step in the process of nitrogen fixation. Nitrogen fixation is required for all forms of life, with nitrogen being essential for the biosynthesis of molecules (nucleotides, amino acids) that create plants, animals and other organisms. They are encoded by the Nif genes or homologs. They are related to protochlorophyllide reductase.
Bioinorganic chemistry is a field that examines the role of metals in biology. Bioinorganic chemistry includes the study of both natural phenomena such as the behavior of metalloproteins as well as artificially introduced metals, including those that are non-essential, in medicine and toxicology. Many biological processes such as respiration depend upon molecules that fall within the realm of inorganic chemistry. The discipline also includes the study of inorganic models or mimics that imitate the behaviour of metalloproteins.

Cysteine dioxygenase (CDO) is a non-heme iron enzyme that catalyzes the conversion of L-cysteine to cysteine sulfinic acid. CDO plays an important role in cysteine catabolism, regulating intracellular levels of cysteine and responding changes in cysteine availability. As such, CDO is highly regulated and undergoes large changes in concentration and efficiency. It oxidizes cysteine to the corresponding sulfinic acid by activation of dioxygen, although the exact mechanism of the reaction is still unclear. In addition to being found in mammals, CDO also exists in some yeast and bacteria, although the exact function is still unknown. CDO has been implicated in various neurodegenerative diseases and cancers, which is likely related to cysteine toxicity.

Xanthinuria, also known as xanthine oxidase deficiency, is a rare genetic disorder causing the accumulation of xanthine. It is caused by a deficiency of the enzyme xanthine oxidase.

Molybdopterins are a class of cofactors found in most molybdenum-containing and all tungsten-containing enzymes. Synonyms for molybdopterin are: MPT and pyranopterin-dithiolate. The nomenclature for this biomolecule can be confusing: Molybdopterin itself contains no molybdenum; rather, this is the name of the ligand that will bind the active metal. After molybdopterin is eventually complexed with molybdenum, the complete ligand is usually called molybdenum cofactor.

Sulfite oxidase is an enzyme in the mitochondria of all eukaryotes, with exception of the yeasts. It oxidizes sulfite to sulfate and, via cytochrome c, transfers the electrons produced to the electron transport chain, allowing generation of ATP in oxidative phosphorylation. This is the last step in the metabolism of sulfur-containing compounds and the sulfate is excreted.

Aldehyde oxidase (AO) is a metabolizing enzyme, located in the cytosolic compartment of tissues in many organisms. AO catalyzes the oxidation of aldehydes into carboxylic acid, and in addition, catalyzes the hydroxylation of some heterocycles. It can also catalyze the oxidation of both cytochrome P450 and monoamine oxidase (MAO) intermediate products. AO plays an important role in the metabolism of several drugs.
Molybdenum cofactor sulfurase is an enzyme that in humans is encoded by the MOCOS gene.
Molybdenum cofactor deficiency is a rare human disease in which the absence of molybdopterin – and consequently its molybdenum complex, commonly called molybdenum cofactor – leads to accumulation of toxic levels of sulphite and neurological damage. Usually this leads to death within months of birth, due to the lack of active sulfite oxidase. Furthermore, a mutational block in molybdenum cofactor biosynthesis causes absence of enzyme activity of xanthine dehydrogenase/oxidase and aldehyde oxidase.
Molybdenum deficiency refers to the clinical consequences of inadequate intake of molybdenum in the diet.
Molybdopterin synthase (EC 2.8.1.12, MPT synthase) is an enzyme required to synthesize molybdopterin (MPT) from precursor Z (now known as cyclic pyranopterin monophosphate). Molydopterin is subsequently complexed with molybdenum to form molybdenum cofactor (MoCo). MPT synthase catalyses the following chemical reaction:
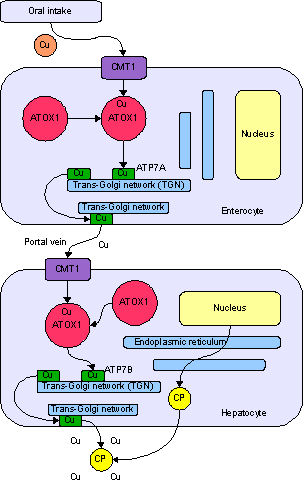
Copper is an essential trace element that is vital to the health of all living things. In humans, copper is essential to the proper functioning of organs and metabolic processes. The human body has complex homeostatic mechanisms which attempt to ensure a constant supply of available copper, while eliminating excess copper whenever this occurs. However, like all essential elements and nutrients, too much or too little nutritional ingestion of copper can result in a corresponding condition of copper excess or deficiency in the body, each of which has its own unique set of adverse health effects.
The aldehyde oxidase and xanthine dehydrogenase, a/b hammerhead domain is an evolutionary conserved protein domain.
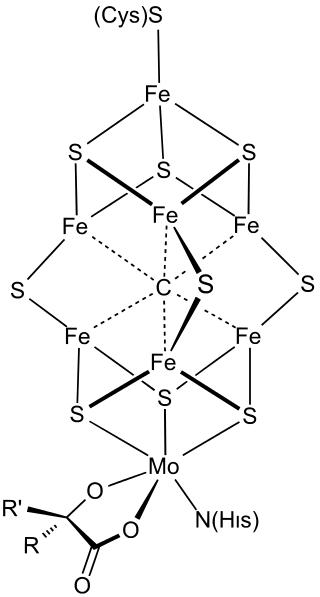
FeMoco (FeMo cofactor) is the primary cofactor of nitrogenase. Nitrogenase is the enzyme that catalyzes the conversion of atmospheric nitrogen molecules N2 into ammonia (NH3) through the process known as nitrogen fixation. Studying FeMoco's role in the reaction mechanism for nitrogen fixation is a potential use case for quantum computers. Even limited quantum computers could enable better simulations of the reaction mechanism. Because it contains iron and molybdenum, the cofactor is called FeMoco. Its stoichiometry is Fe7MoS9C.

Fosdenopterin, sold under the brand name Nulibry, is a medication used to reduce the risk of death due to a rare genetic disease known as molybdenum cofactor deficiency type A.
{{cite book}}: CS1 maint: DOI inactive as of January 2024 (link) electronic-book ISBN 978-94-007-5561-1 ISSN 1559-0836 electronic- ISSN 1868-0402 {{cite journal}}: CS1 maint: DOI inactive as of February 2024 (link)