The dicotyledons, also known as dicots, are one of the two groups into which all the flowering plants (angiosperms) were formerly divided. The name refers to one of the typical characteristics of the group: namely, that the seed has two embryonic leaves or cotyledons. There are around 200,000 species within this group. The other group of flowering plants were called monocotyledons, typically each having one cotyledon. Historically, these two groups formed the two divisions of the flowering plants.

Monocotyledons, commonly referred to as monocots, are grass and grass-like flowering plants (angiosperms), the seeds of which typically contain only one embryonic leaf, or cotyledon. They constitute one of the major groups into which the flowering plants have traditionally been divided; the rest of the flowering plants have two cotyledons and are classified as dicotyledons, or dicots.
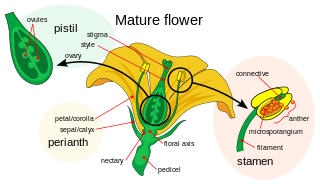
Petals are modified leaves that surround the reproductive parts of flowers. They are often brightly colored or unusually shaped to attract pollinators. All of the petals of a flower are collectively known as the corolla. Petals are usually accompanied by another set of modified leaves called sepals, that collectively form the calyx and lie just beneath the corolla. The calyx and the corolla together make up the perianth, the non-reproductive portion of a flower. When the petals and sepals of a flower are difficult to distinguish, they are collectively called tepals. Examples of plants in which the term tepal is appropriate include genera such as Aloe and Tulipa. Conversely, genera such as Rosa and Phaseolus have well-distinguished sepals and petals. When the undifferentiated tepals resemble petals, they are referred to as "petaloid", as in petaloid monocots, orders of monocots with brightly colored tepals. Since they include Liliales, an alternative name is lilioid monocots.

Pollination is the transfer of pollen from an anther of a plant to the stigma of a plant, later enabling fertilisation and the production of seeds. Pollinating agents can be animals such as insects, for example beetles or butterflies; birds, and bats; water; wind; and even plants themselves. Pollinating animals travel from plant to plant carrying pollen on their bodies in a vital interaction that allows the transfer of genetic material critical to the reproductive system of most flowering plants. When self-pollination occurs within a closed flower. Pollination often occurs within a species. When pollination occurs between species, it can produce hybrid offspring in nature and in plant breeding work.

Iridaceae is a family of plants in order Asparagales, taking its name from the irises. It has a nearly global distribution, with 69 accepted genera with a total of c. 2500 species. It includes a number of economically important cultivated plants, such as species of Freesia, Gladiolus, and Crocus, as well as the crop saffron.

Self-pollination is a form of pollination in which pollen from the same plant arrives at the stigma of a flower or at the ovule. There are two types of self-pollination: in autogamy, pollen is transferred to the stigma of the same flower; in geitonogamy, pollen is transferred from the anther of one flower to the stigma of another flower on the same flowering plant, or from microsporangium to ovule within a single (monoecious) gymnosperm. Some plants have mechanisms that ensure autogamy, such as flowers that do not open (cleistogamy), or stamens that move to come into contact with the stigma. The term selfing that is often used as a synonym, is not limited to self-pollination, but also applies to other type of self-fertilization.

Plant reproductive morphology is the study of the physical form and structure of those parts of plants directly or indirectly concerned with sexual reproduction.
Self-incompatibility (SI) is a general name for several genetic mechanisms that prevent self-fertilization in sexually reproducing organisms, and thus encourage outcrossing and allogamy. It is contrasted with separation of sexes among individuals (dioecy), and their various modes of spatial (herkogamy) and temporal (dichogamy) separation.

Heterostyly is a unique form of polymorphism and herkogamy in flowers. In a heterostylous species, two or three morphological types of flowers, termed "morphs", exist in the population. On each individual plant, all flowers share the same morph. The flower morphs differ in the lengths of the pistil and stamens, and these traits are not continuous. The morph phenotype is genetically linked to genes responsible for a unique system of self-incompatibility, termed heteromorphic self-incompatibility, that is, the pollen from a flower on one morph cannot fertilize another flower of the same morph.

Entomophily or insect pollination is a form of pollination whereby pollen of plants, especially but not only of flowering plants, is distributed by insects. Flowers pollinated by insects typically advertise themselves with bright colours, sometimes with conspicuous patterns leading to rewards of pollen and nectar; they may also have an attractive scent which in some cases mimics insect pheromones. Insect pollinators such as bees have adaptations for their role, such as lapping or sucking mouthparts to take in nectar, and in some species also pollen baskets on their hind legs. This required the coevolution of insects and flowering plants in the development of pollination behaviour by the insects and pollination mechanisms by the flowers, benefiting both groups. Both the size and the density of a population are known to affect pollination and subsequent reproductive performance.

The stigma is the receptive tip of a carpel, or of several fused carpels, in the gynoecium of a flower.

A flower, also known as a bloom or blossom, is the reproductive structure found in flowering plants. Flowers consist of a combination of vegetative organs – sepals that enclose and protect the developing flower, petals that attract pollinators, and reproductive organs that produce gametophytes, which in flowering plants produce gametes. The male gametophytes, which produce sperm, are enclosed within pollen grains produced in the anthers. The female gametophytes are contained within the ovules produced in the carpels.

Cymodoceaceae is a family of flowering plants, sometimes known as the "manatee-grass family", which includes only marine species.

Ornithophily or bird pollination is the pollination of flowering plants by birds. This sometimes coevolutionary association is derived from insect pollination (entomophily) and is particularly well developed in some parts of the world, especially in the tropics, Southern Africa, and on some island chains. The association involves several distinctive plant adaptations forming a "pollination syndrome". The plants typically have colourful, often red, flowers with long tubular structures holding ample nectar and orientations of the stamen and stigma that ensure contact with the pollinator. Birds involved in ornithophily tend to be specialist nectarivores with brushy tongues and long bills, that are either capable of hovering flight or light enough to perch on the flower structures.

Pollination syndromes are suites of flower traits that have evolved in response to natural selection imposed by different pollen vectors, which can be abiotic or biotic, such as birds, bees, flies, and so forth through a process called pollinator-mediated selection. These traits include flower shape, size, colour, odour, reward type and amount, nectar composition, timing of flowering, etc. For example, tubular red flowers with copious nectar often attract birds; foul smelling flowers attract carrion flies or beetles, etc.
Plant reproduction is the production of new offspring in plants, which can be accomplished by sexual or asexual reproduction. Sexual reproduction produces offspring by the fusion of gametes, resulting in offspring genetically different from either parent. Asexual reproduction produces new individuals without the fusion of gametes, resulting in clonal plants that are genetically identical to the parent plant and each other, unless mutations occur.
Sexual selection is described as natural selection arising through preference by one sex for certain characteristics in individuals of the other sex. Sexual selection is a common concept in animal evolution but, with plants, it is oftentimes overlooked because many plants are hermaphrodites. Flowering plants show many characteristics that are often sexually selected for. For example, flower symmetry, nectar production, floral structure, and inflorescences are just a few of the many secondary sex characteristics acted upon by sexual selection. Sexual dimorphisms and reproductive organs can also be affected by sexual selection in flowering plants.
Reproductive assurance occurs as plants have mechanisms to assure full seed set through selfing when outcross pollen is limiting. It is assumed that self-pollination is beneficial, in spite of potential fitness costs, when there is insufficient pollinator services or outcross pollen from other individuals to accomplish full seed set.. This phenomenon has been observed since the 19th century, when Darwin observed that self-pollination was common in some plants. Constant pollen limitation may cause the evolution of automatic selfing, also known as autogamy. This occurs in plants such as weeds, and is a form of reproductive assurance. As plants pursue reproductive assurance through self-fertilization, there is an increase in homozygosity, and inbreeding depression, due to genetic load, which results in reduced fitness of selfed offspring. Solely outcrossing plants may not be successful colonizers of new regions due to lack of other plants to outcross with, so colonizing species are expected to have mechanisms of reproductive assurance - an idea first proposed by Herbert G. Baker and referred to as Baker's "law" or "rule". Baker's law predicts that reproductive assurance affects establishment of plants in many contexts, including spread by weedy plants and following long-distance dispersal, such as occurs during island colonization. As plants evolve towards increase self-fertilization, energy is redirected to seed production rather than characteristics that increased outcrossing, such as floral attractants, which is a condition known as the selfing syndrome.
Cryptic self-incompatibility (CSI) is the botanical expression that's used to describe a weakened self-incompatibility (SI) system. CSI is one expression of a mixed mating system in flowering plants. Both SI and CSI are traits that increase the frequency of fertilization of ovules by outcross pollen, as opposed to self-pollen.

The pollination of orchids is a complex chapter in the biology of this family of plants that are distinguished by the complexity of their flowers and by intricate ecological interactions with their pollinator agents. It has captured the attention of numerous scientists over time, including Charles Darwin, father of the theory of evolution by natural selection. Darwin published in 1862 the first observations of the fundamental role of insects in orchid pollination, in his book The Fertilization of Orchids. Darwin stated that the varied stratagems orchids use to attract their pollinators transcend the imagination of any human being.