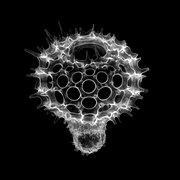
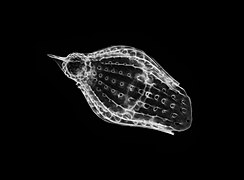
The Acantharea (Acantharia) are a group of radiolarian protozoa, distinguished mainly by their strontium sulfate skeletons. Acantharians are heterotrophic marine microplankton that range in size from about 200 microns in diameter up to several millimeters. Some acantharians have photosynthetic endosymbionts and hence are considered mixotrophs.
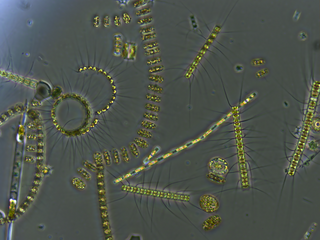
Zooplankton are the animal component of the planktonic community. Plankton are aquatic organisms that are unable to swim effectively against currents. Consequently, they drift or are carried along by currents in the ocean, or by currents in seas, lakes or rivers.

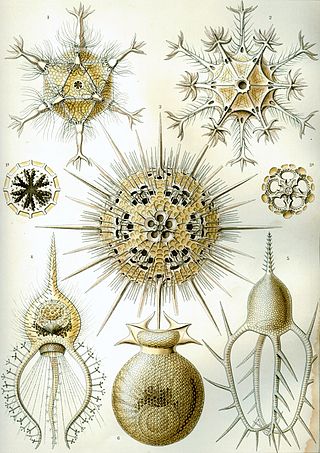
The Radiolaria, also called Radiozoa, are protozoa of diameter 0.1–0.2 mm that produce intricate mineral skeletons, typically with a central capsule dividing the cell into the inner and outer portions of endoplasm and ectoplasm. The elaborate mineral skeleton is usually made of silica. They are found as zooplankton throughout the global ocean. As zooplankton, radiolarians are primarily heterotrophic, but many have photosynthetic endosymbionts and are, therefore, considered mixotrophs. The skeletal remains of some types of radiolarians make up a large part of the cover of the ocean floor as siliceous ooze. Due to their rapid change as species and intricate skeletons, radiolarians represent an important diagnostic fossil found from the Cambrian onwards.

The polycystines are a group of radiolarians. They include the vast majority of the fossil radiolaria, as their skeletons are abundant in marine sediments, making them one of the most common groups of microfossils. These skeletons are composed of opaline silica. In some it takes the form of relatively simple spicules, but in others it forms more elaborate lattices, such as concentric spheres with radial spines or sequences of conical chambers. Two of the orders belonging to this group are the radially-symmetrical Spumellaria, dating back to the late Cambrian period, and the bilaterally-symmetrical Nasselaria, whose origin is placed within the lower Devonian.

Micropaleontology is the branch of paleontology (palaeontology) that studies microfossils, or fossils that require the use of a microscope to see the organism, its morphology and its characteristic details.
Phaeodarea or Phaeodaria is a group of amoeboid cercozoan organisms. They are traditionally considered radiolarians, but in molecular trees do not appear to be close relatives of the other groups, and are instead placed among the Cercozoa. They are distinguished by the structure of their central capsule and by the presence of a phaeodium, an aggregate of waste particles within the cell.

A microfossil is a fossil that is generally between 0.001 mm and 1 mm in size, the visual study of which requires the use of light or electron microscopy. A fossil which can be studied with the naked eye or low-powered magnification, such as a hand lens, is referred to as a macrofossil.
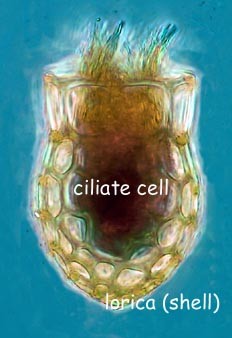
Tintinnids are ciliates of the choreotrich order Tintinnida, distinguished by vase-shaped shells, the name deriving from a Latin source meaning a small tinkling bell, that are called loricae, which are mostly protein but may incorporate minute pieces of minerals.
A protist or protoctist is any eukaryotic organism that is not an animal, plant, or fungus. Protists do not form a natural group, or clade, but an artificial grouping of several independent clades that evolved from the last eukaryotic common ancestor.
Syndinium is a cosmopolitan genus of parasitic dinoflagellates that infest and kill marine planktonic species of copepods and radiolarians. Syndinium belongs to order Syndiniales, a candidate for the uncultured group I and II marine alveolates. The lifecycle of Syndinium is not well understood beyond the parasitic and zoospore stages.

Spumellaria is an order of radiolarians in the class Polycystinea. They are ameboid protists appearing in abundance in the world's oceans, possessing a radially-symmetrical silica (opal) skeleton that has ensured their preservation in fossil records. They belong among the oldest Polycystine organisms, dating back to the lower Cambrian. Historically, many concentric radiolarians have been included in the Spumellaria order based on the absence of the initial spicular system, an early-develop structure that, by its lacking, sets them apart from Entactinaria despite their similar morphology. Living exemplars of the order feed by catching prey, such as copepod nauplii or tintinnids, on the adhesive ends of their pseudopodia extending radially from their skeleton; however, some have been observed as mixotrophs living in symbiosis with various photosynthetic algal organisms such as dinoflagellates, cyanobacteria, prasinophytes or haptophytes, which may cause their distribution to center in the greatest abundance and diversity within trophical waters.

Marine microorganisms are defined by their habitat as microorganisms living in a marine environment, that is, in the saltwater of a sea or ocean or the brackish water of a coastal estuary. A microorganism is any microscopic living organism or virus, that is too small to see with the unaided human eye without magnification. Microorganisms are very diverse. They can be single-celled or multicellular and include bacteria, archaea, viruses and most protozoa, as well as some fungi, algae, and animals, such as rotifers and copepods. Many macroscopic animals and plants have microscopic juvenile stages. Some microbiologists also classify viruses as microorganisms, but others consider these as non-living.
Vitrella brassicaformis (CCMP3155) is a unicellular alga belonging to the eukaryotic supergroup Alveolata. V. brassicaformis and its closest known relative, Chromera velia, are the only two currently described members of the phylum Chromerida, which in turn constitutes part of the taxonomically unranked group Colpodellida. Chromerida is phylogenetically closely related to the phylum Apicomplexa, which includes Plasmodium, the agent of malaria. Notably, both V. brassicaformis and C. velia are photosynthetic, each containing a complex secondary plastid. This characteristic defined the discovery of these so-called 'chromerids,' as their photosynthetic capacity positioned them to shed light upon the evolution of Apicomplexa's non-photosynthetic parasitism. Both genera lack chlorophyll b or c; these absences link the two taxonomically, as algae bearing only chlorophyll a are rare amid the biodiversity of life. Despite their similarities, V. brassicaformis differs significantly from C. velia in morphology, lifecycle, and accessory photosynthetic pigmentation. V. brassicaformis has a green color, with a complex lifecycle involving multiple pathways and a range of sizes and morphologies, while Chromera has a brown color and cycles through a simpler process from generation to generation. The color differences are due to differences in accessory pigments.

Collodaria is a unicellular order under the phylum Radiozoa and the infrakingdom Rhizaria. Like most of the Radiolaria taxonomy, Collodaria was first described by Ernst Haeckel, a German scholar who published three volumes of manuscript describing the extensive samples of Radiolaria collected by the voyage of HMS Challenger. Recent molecular phylogenetic studies concluded that there are Collodaria contains three families, Sphaerozodae, Collosphaeridae, and Collophidilidae.
Pelagodinium béii is a photosynthetic dinoflagellate that forms a symbiotic relationship with planktonic foraminifera.
The genus Stylodictya belongs to a group of organisms called the Radiolaria. Radiolarians are amoeboid protists found as zooplankton in oceans around the world and are typically identified by their ornate skeletons.

Cornutella profunda is a species of radiolarian in the family Theoperidae and the genus Cornutella. The abundance and actual geographic span of C. profunda has not yet been fully explored, however few have been caught in various regions around the world. Samples have seen in larger numbers in the Adriatic Sea, the South China Sea, and far off the coast of Southern Africa near Namibia, and in smaller numbers in all other oceans around the world.

Marine protists are defined by their habitat as protists that live in marine environments, that is, in the saltwater of seas or oceans or the brackish water of coastal estuaries. Life originated as marine single-celled prokaryotes and later evolved into more complex eukaryotes. Eukaryotes are the more developed life forms known as plants, animals, fungi and protists. Protists are the eukaryotes that cannot be classified as plants, fungi or animals. They are mostly single-celled and microscopic. The term protist came into use historically as a term of convenience for eukaryotes that cannot be strictly classified as plants, animals or fungi. They are not a part of modern cladistics because they are paraphyletic.

Many protists have protective shells or tests, usually made from silica (glass) or calcium carbonate (chalk). Protists are a diverse group of eukaryote organisms that are not plants, animals, or fungi. They are typically microscopic unicellular organisms that live in water or moist environments.
A protist is any eukaryotic organism that is not an animal, plant, or fungus. While it is likely that protists share a common ancestor, the last eukaryotic common ancestor, the exclusion of other eukaryotes means that protists do not form a natural group, or clade. Therefore, some protists may be more closely related to animals, plants, or fungi than they are to other protists. However, like algae, invertebrates and protozoans, the grouping is used for convenience.