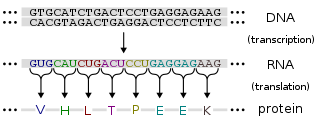
A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, its root system. Mycorrhizae play important roles in plant nutrition, soil biology and soil chemistry.

Medicago truncatula, the barrelclover, strong-spined medick, barrel medic, or barrel medick, is a small annual legume native to the Mediterranean region that is used in genomic research. It is a low-growing, clover-like plant 10–60 centimetres (3.9–23.6 in) tall with trifoliate leaves. Each leaflet is rounded, 1–2 centimetres (0.39–0.79 in) long, often with a dark spot in the center. The flowers are yellow, produced singly or in a small inflorescence of two to five together; the fruit is a small, spiny pod.
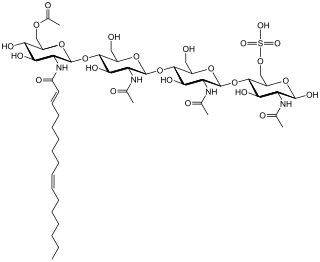
Nod factors, are signaling molecules produced by soil bacteria known as rhizobia during the initiation of nodules on the root of legumes. A symbiosis is formed when legumes take up the bacteria. The rhizobia produce nitrogen for the plant, and the legumes produce leghemoglobin to carry away any oxygen that would inhibit nitrogenase activity.

Primary succession is one of two types of biological and ecological succession of plant life, occurring in an environment in which new substrate devoid of vegetation and other organisms usually lacking soil, such as a lava flow or area left from retreated glacier, is deposited. In other words, it is the gradual growth of an ecosystem over a longer period of time.
Glomeromycota are one of eight currently recognized divisions within the kingdom Fungi, with approximately 230 described species. Members of the Glomeromycota form arbuscular mycorrhizas (AMs) with the thalli of bryophytes and the roots of vascular land plants. Not all species have been shown to form AMs, and one, Geosiphon pyriformis, is known not to do so. Instead, it forms an endocytobiotic association with Nostoc cyanobacteria. The majority of evidence shows that the Glomeromycota are dependent on land plants for carbon and energy, but there is recent circumstantial evidence that some species may be able to lead an independent existence. The arbuscular mycorrhizal species are terrestrial and widely distributed in soils worldwide where they form symbioses with the roots of the majority of plant species (>80%). They can also be found in wetlands, including salt-marshes, and associated with epiphytic plants.
Glomus is a genus of arbuscular mycorrhizal (AM) fungi, and all species form symbiotic relationships (mycorrhizas) with plant roots. Glomus is the largest genus of AM fungi, with ca. 85 species described, but is currently defined as non-monophyletic.

The ericoid mycorrhiza is a mutualistic relationship formed between members of the plant family Ericaceae and several lineages of mycorrhizal fungi. This symbiosis represents an important adaptation to acidic and nutrient poor soils that species in the Ericaceae typically inhabit, including boreal forests, bogs, and heathlands. Molecular clock estimates suggest that the symbiosis originated approximately 140 million years ago.
Microbial inoculants also known as soil inoculants are agricultural amendments that use beneficial endophytes (microbes) to promote plant health. Many of the microbes involved form symbiotic relationships with the target crops where both parties benefit (mutualism). While microbial inoculants are applied to improve plant nutrition, they can also be used to promote plant growth by stimulating plant hormone production.
The mycorrhizosphere is the region around a mycorrhizal fungus in which nutrients released from the fungus increase the microbial population and its activities. The roots of most terrestrial plants, including most crop plants and almost all woody plants, are colonized by mycorrhiza-forming symbiotic fungi. In this relationship, the plant roots are infected by a fungus, but the rest of the fungal mycelium continues to grow through the soil, digesting and absorbing nutrients and water and sharing these with its plant host. The fungus in turn benefits by receiving photosynthetic sugars from its host. The mycorrhizosphere consists of roots, hyphae of the directly connected mycorrhizal fungi, associated microorganisms, and the soil in their direct influence.
Mycorrhizal networks are underground hyphal networks created by mycorrhizal fungi that connect individual plants together and transfer water, carbon, nitrogen, and other nutrients and minerals. The formation of these networks is context dependent, and can be influenced by factors such as soil fertility, resource availability, host or myco-symbiont genotype, disturbance and seasonal variation. By analogy to the many roles intermediated by the World Wide Web in human communities, the many roles that mycorrhizal networks appear to play in woodland have earned them a colloquial nickname: the Wood Wide Web.. Wood wide web can be described as plants’ way of talking. Plants communicate through the mycorrhizal network in a similar manner to thoughts travelling over the brain’s network of neurons.

Soil carbon storage is an important function of terrestrial ecosystems. Soil contains more carbon than plants and the atmosphere combined. Understanding what maintains the soil carbon pool is important to understand the current distribution of carbon on Earth, and how it will respond to environmental change. While much research has been done on how plants, free-living microbial decomposers, and soil minerals affect this pool of carbon, it is recently coming to light that mycorrhizal fungi—symbiotic fungi that associate with roots of almost all living plants—may play an important role in maintaining this pool as well. Measurements of plant carbon allocation to mycorrhizal fungi have been estimated to be 5-20% of total plant carbon uptake, and in some ecosystems the biomass of mycorrhizal fungi can be comparable to the biomass of fine roots. Recent research has shown that mycorrhizal fungi hold 50 to 70 percent of the total carbon stored in leaf litter and soil on forested islands in Sweden. Turnover of mycorrhizal biomass into the soil carbon pool is thought to be rapid and has been shown in some ecosystems to be the dominant pathway by which living carbon enters the soil carbon pool.

An ectomycorrhiza is a form of symbiotic relationship that occurs between a fungal symbiont and the roots of various plant species. The mycobiont tends to be predominantly from the phyla Basidiomycota and Ascomycota, although a few are represented in the phylum Zygomycota. Ectomycorrhizas form between fungi and the roots of around 2% of plant species. These tend to be composed of woody plants, including species from the birch, dipterocarp, myrtle, beech, willow, pine and rose families.

Rhizophagus is a genus of arbuscular mycorrhizal (AM) fungi that form symbiotic relationships (mycorrhizas) with plant roots. The genome of Rhizophagus irregularis was recently sequenced.

Rhizophagus irregularis is an arbuscular mycorrhizal fungus used as a soil inoculant in agriculture and horticulture. In addition, it is one of the best mycorrhizal varieties of fungi available to mycoforestry, but as it does not produce fruiting bodies it "has virtually no market value as an edible or medicinal mushroom"
Dark septate endophytes (DSE) are a group of endophytic fungi characterized by their morphology of melanized, septate, hyphae. This group is likely paraphyletic, and contain conidial as well as sterile fungi that colonize roots intracellularly or intercellularly. Very little is known about the number of fungal taxa within this group, but all are in the Ascomycota. They are found in over 600 plant species and across 114 families of angiosperms and gymnosperms and co-occur with other types of mycorrhizal fungi. They have a wide global distribution and can be more abundant in stressed environments. Much of their taxonomy, physiology, and ecology are unknown.
Orchid mycorrhizae are symbiotic relationships between the roots of plants of the family Orchidaceae and a variety of fungi. All orchids are myco-heterotrophic at some point in their life cycle. Orchid mycorrhizae are critically important during orchid germination, as an orchid seed has virtually no energy reserve and obtains its carbon from the fungal symbiont.
Mycorrhiza helper bacteria (MHB) are a group of organisms that form symbiotic associations with both ectomycorrhiza and arbuscular mycorrhiza. MHBs are diverse and belong to different bacterial phyla including both gram-negative and gram-positive bacteria. Some of the most common types are Pseudomonas and Streptomyces. MHBs have specific interactions with fungi, but not with the plants. MHB enhance mycorrhizal function, increase mycorrhizal growth, provide nutrients to the fungus and plant, improve soil conductance, select to aid pathogens, and help promote defense mechanisms.
Plant to plant communication via mycorrhizal networks refers to connections through mycorrhizal networks that facilitate communication between plants of the same or different species. Mycorrhizal networks allow for the transfers of signals and cues between plants which influence the behavior of the connected plants by inducing morphological or physiological changes. The chemical substances which act as these signals and cues are referred to as infochemicals. These can be allelochemicals, defensive chemicals or nutrients. Allelochemicals are used by plants to interfere with the growth or development of other plants or organisms, defensive chemicals can help plants in mycorrhizal networks defend themselves against attack by pathogens or herbivores, and transferred nutrients can affect growth and nutrition. Results of studies which demonstrate these modes of communication have led the authors to hypothesize mechanisms by which the transfer of these nutrients can affect the fitness of the connected plants.
Mycorrhizae and changing climate refers to the effects of changing climates on mycorrhizae, the symbiotic association between a fungus and the roots of a vascular host plant.