
Cyanobacteria, also called Cyanobacteriota or Cyanophyta, are a phylum of gram-negative bacteria that obtain energy via photosynthesis. The name cyanobacteria refers to their color, which similarly forms the basis of cyanobacteria's common name, blue-green algae, although they are not usually scientifically classified as algae. They appear to have originated in a freshwater or terrestrial environment. Cyanobacteria produce a range of toxins known as cyanotoxins that can cause harmful health effects in humans and animals.

Xanthophylls are yellow pigments that occur widely in nature and form one of two major divisions of the carotenoid group; the other division is formed by the carotenes. The name is from Greek: xanthos (ξανθός), meaning "yellow", and phyllon (φύλλον), meaning "leaf"), due to their formation of the yellow band seen in early chromatography of leaf pigments.

Phycocyanin is a pigment-protein complex from the light-harvesting phycobiliprotein family, along with allophycocyanin and phycoerythrin. It is an accessory pigment to chlorophyll. All phycobiliproteins are water-soluble, so they cannot exist within the membrane like carotenoids can. Instead, phycobiliproteins aggregate to form clusters that adhere to the membrane called phycobilisomes. Phycocyanin is a characteristic light blue color, absorbing orange and red light, particularly near 620 nm, and emits fluorescence at about 650 nm. Allophycocyanin absorbs and emits at longer wavelengths than phycocyanin C or phycocyanin R. Phycocyanins are found in cyanobacteria. Phycobiliproteins have fluorescent properties that are used in immunoassay kits. Phycocyanin is from the Greek phyco meaning “algae” and cyanin is from the English word “cyan", which conventionally means a shade of blue-green and is derived from the Greek “kyanos" which means a somewhat different color: "dark blue". The product phycocyanin, produced by Aphanizomenon flos-aquae and Spirulina, is for example used in the food and beverage industry as the natural coloring agent 'Lina Blue' or 'EXBERRY Shade Blue' and is found in sweets and ice cream. In addition, fluorescence detection of phycocyanin pigments in water samples is a useful method to monitor cyanobacteria biomass.

Phycobilisomes are light harvesting antennae of photosystem II in cyanobacteria, red algae and glaucophytes. It was lost in the plastids of green algae / plants (chloroplasts).

Carboxysomes are bacterial microcompartments (BMCs) consisting of polyhedral protein shells filled with the enzymes ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO)—the predominant enzyme in carbon fixation and the rate limiting enzyme in the Calvin cycle—and carbonic anhydrase.

Allophycocyanin is a protein from the light-harvesting phycobiliprotein family, along with phycocyanin, phycoerythrin and phycoerythrocyanin. It is an accessory pigment to chlorophyll. All phycobiliproteins are water-soluble and therefore cannot exist within the membrane like carotenoids, but aggregate, forming clusters that adhere to the membrane called phycobilisomes. Allophycocyanin absorbs and emits red light, and is readily found in Cyanobacteria, and red algae. Phycobilin pigments have fluorescent properties that are used in immunoassay kits. In flow cytometry, it is often abbreviated APC. To be effectively used in applications such as FACS, High-Throughput Screening (HTS) and microscopy, APC needs to be chemically cross-linked.
Phycobiliproteins are water-soluble proteins present in cyanobacteria and certain algae. They capture light energy, which is then passed on to chlorophylls during photosynthesis. Phycobiliproteins are formed of a complex between proteins and covalently bound phycobilins that act as chromophores. They are most important constituents of the phycobilisomes.
A light-harvesting complex consists of a number of chromophores which are complex subunit proteins that may be part of a larger super complex of a photosystem, the functional unit in photosynthesis. It is used by plants and photosynthetic bacteria to collect more of the incoming light than would be captured by the photosynthetic reaction center alone. The light which is captured by the chromophores is capable of exciting molecules from their ground state to a higher energy state, known as the excited state. This excited state does not last very long and is known to be short-lived.
Photoprotection is the biochemical process that helps organisms cope with molecular damage caused by sunlight. Plants and other oxygenic phototrophs have developed a suite of photoprotective mechanisms to prevent photoinhibition and oxidative stress caused by excess or fluctuating light conditions. Humans and other animals have also developed photoprotective mechanisms to avoid UV photodamage to the skin, prevent DNA damage, and minimize the downstream effects of oxidative stress.
In biophysics, the Kautsky effect is a phenomenon consisting of a typical variation in the behavior of a plant fluorescence when exposed to light. It was discovered in 1931 by H. Kautsky and A. Hirsch.

Photoinhibition is light-induced reduction in the photosynthetic capacity of a plant, alga, or cyanobacterium. Photosystem II (PSII) is more sensitive to light than the rest of the photosynthetic machinery, and most researchers define the term as light-induced damage to PSII. In living organisms, photoinhibited PSII centres are continuously repaired via degradation and synthesis of the D1 protein of the photosynthetic reaction center of PSII. Photoinhibition is also used in a wider sense, as dynamic photoinhibition, to describe all reactions that decrease the efficiency of photosynthesis when plants are exposed to light.
Synechocystis sp. PCC6803 is a strain of unicellular, freshwater cyanobacteria. Synechocystis sp. PCC6803 is capable of both phototrophic growth by oxygenic photosynthesis during light periods and heterotrophic growth by glycolysis and oxidative phosphorylation during dark periods. Gene expression is regulated by a circadian clock and the organism can effectively anticipate transitions between the light and dark phases.
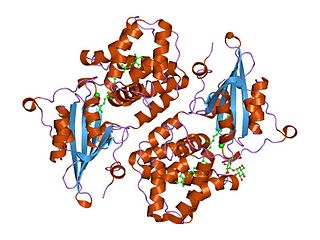
In molecular biology the orange carotenoid N-terminal domain is a protein domain found predominantly at the N-terminus of the Orange carotenoid protein (OCP), and is involved in non-covalent binding of a carotenoid chromophore. It is unique for being present in soluble proteins, whereas the vast majority of domains capable of binding carotenoids are intrinsic membrane proteins. Thus far, it has exclusively been found in cyanobacteria, among which it is widespread. The domain also exists on its own, in uncharacterized cyanobacterial proteins referred to as "Red Carotenoid Protein" (RCP). The domain adopts an alpha-helical structure consisting of two four-helix bundles.
Antheraxanthin is a bright yellow accessory pigment found in many organisms that perform photosynthesis. It is a xanthophyll cycle pigment, an oil-soluble alcohol within the xanthophyll subgroup of carotenoids. Antheraxanthin is both a component in and product of the cellular photoprotection mechanisms in photosynthetic green algae, red algae, euglenoids, and plants.
Iron-starvation-induced protein A, also known as isiA, is a photosynthesis-related chlorophyll-containing protein found in cyanobacteria. It belongs to the chlorophyll-a/b-binding family of proteins, and has been shown to have a photoprotection role in preventing oxidative damage via energy dissipation. It was originally identified under Fe starvation, and thus received the name iron-starvation-induced protein A. However, the protein has more recently been found to respond to a variety of stress conditions such as high irradiance. It can aggregate with carotenoids and form rings around the PSI reaction center complexes to aid in photoprotective energy dissipation.
In photosynthesis, state transitions are rearrangements of the photosynthetic apparatus which occur on short time-scales. The effect is prominent in cyanobacteria, whereby the phycobilisome light-harvesting antenna complexes alter their preference for transfer of excitation energy between the two reaction centers, PS I and PS II. This shift helps to minimize photodamage caused by reactive oxygen species (ROS) under stressful conditions such as high light, but may also be used to offset imbalances between the rates of generating reductant and ATP.
3′-Hydroxyechinenone is a keto-carotenoid pigment found in cyanobacteria and microalgae. Carotenoids belong to a larger class of phytochemicals known as terpenoids. The chemical formula of canthaxanthin is C40H54O2. It is found non-covalently bound in the orange carotenoid protein (OCP), which is a soluble protein involved in photoprotection and non-photochemical quenching of photosynthesis.
Fluorescence recovery protein (FRP) is a small protein involved in regulating non-photochemical quenching in cyanobacteria. It prevents accumulation of the red photoactivated form of orange carotenoid protein (OCP), thereby reducing the amount of fluorescence quenching that occurs between the OCP and the phycobilisome antenna complexes. It interacts with the C-terminal domain of OCP, which shares homology with the NTF2 superfamily.
David W. Krogmann was an American biologist and a professor of biochemistry at Purdue University. He is known for his work in photosynthesis in chloroplasts and cyanobacteria.
In molecular biology, the PYP domain is a p-coumaric acid-binding protein domain. They are present in various proteins in bacteria.


