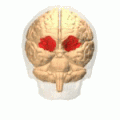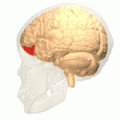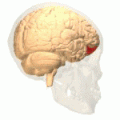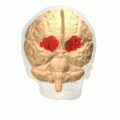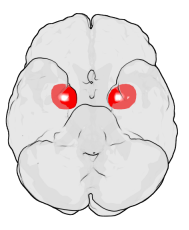
The amygdala is one of two almond-shaped clusters of nuclei located deep and medially within the temporal lobes of the brain's cerebrum in complex vertebrates, including humans. Shown to perform a primary role in the processing of memory, decision making, and emotional responses, the amygdalae are considered part of the limbic system. The term "amygdala" was first introduced by Karl Friedrich Burdach in 1822.
The mesolimbic pathway, sometimes referred to as the reward pathway, is a dopaminergic pathway in the brain. The pathway connects the ventral tegmental area in the midbrain to the ventral striatum of the basal ganglia in the forebrain. The ventral striatum includes the nucleus accumbens and the olfactory tubercle.

The cingulate cortex is a part of the brain situated in the medial aspect of the cerebral cortex. The cingulate cortex includes the entire cingulate gyrus, which lies immediately above the corpus callosum, and the continuation of this in the cingulate sulcus. The cingulate cortex is usually considered part of the limbic lobe.

The caudate nucleus is one of the structures that make up the corpus striatum, which is a component of the basal ganglia in the human brain. While the caudate nucleus has long been associated with motor processes due to its role in Parkinson's disease, it plays important roles in various other nonmotor functions as well, including procedural learning, associative learning and inhibitory control of action, among other functions. The caudate is also one of the brain structures which compose the reward system and functions as part of the cortico–basal ganglia–thalamic loop.

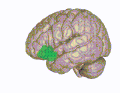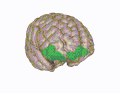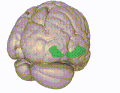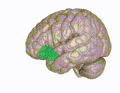
The olfactory bulb is a neural structure of the vertebrate forebrain involved in olfaction, the sense of smell. It sends olfactory information to be further processed in the amygdala, the orbitofrontal cortex (OFC) and the hippocampus where it plays a role in emotion, memory and learning. The bulb is divided into two distinct structures: the main olfactory bulb and the accessory olfactory bulb. The main olfactory bulb connects to the amygdala via the piriform cortex of the primary olfactory cortex and directly projects from the main olfactory bulb to specific amygdala areas. The accessory olfactory bulb resides on the dorsal-posterior region of the main olfactory bulb and forms a parallel pathway. Destruction of the olfactory bulb results in ipsilateral anosmia, while irritative lesions of the uncus can result in olfactory and gustatory hallucinations.

The frontal lobe is the largest of the four major lobes of the brain in mammals, and is located at the front of each cerebral hemisphere. It is parted from the parietal lobe by a groove between tissues called the central sulcus and from the temporal lobe by a deeper groove called the lateral sulcus. The most anterior rounded part of the frontal lobe is known as the frontal pole, one of the three poles of the cerebrum.

Brodmann area 10 is the anterior-most portion of the prefrontal cortex in the human brain. BA10 was originally defined broadly in terms of its cytoarchitectonic traits as they were observed in the brains of cadavers, but because modern functional imaging cannot precisely identify these boundaries, the terms anterior prefrontal cortex, rostral prefrontal cortex and frontopolar prefrontal cortex are used to refer to the area in the most anterior part of the frontal cortex that approximately covers BA10—simply to emphasize the fact that BA10 does not include all parts of the prefrontal cortex.

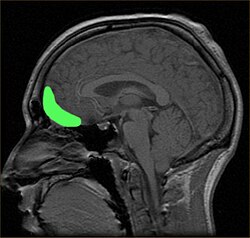
Brodmann area 11 is one of Brodmann's cytologically defined regions of the brain. It is in the orbitofrontal cortex which is above the eye sockets (orbitae). It is involved in decision making, processing rewards, and encoding new information into long-term memory..

Dopaminergic pathways in the human brain are involved in both physiological and behavioral processes including movement, cognition, executive functions, reward, motivation, and neuroendocrine control. Each pathway is a set of projection neurons, consisting of individual dopaminergic neurons.

The ventral tegmental area (VTA), also known as the ventral tegmental area of Tsai, or simply ventral tegmentum, is a group of neurons located close to the midline on the floor of the midbrain. The VTA is the origin of the dopaminergic cell bodies of the mesocorticolimbic dopamine system and other dopamine pathways; it is widely implicated in the drug and natural reward circuitry of the brain. The VTA plays an important role in a number of processes, including reward cognition and orgasm, among others, as well as several psychiatric disorders. Neurons in the VTA project to numerous areas of the brain, ranging from the prefrontal cortex to the caudal brainstem and several regions in between.
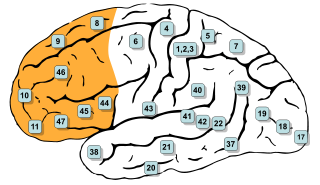
In mammalian brain anatomy, the prefrontal cortex (PFC) covers the front part of the frontal lobe of the cerebral cortex. The PFC contains the Brodmann areas BA8, BA9, BA10, BA11, BA12, BA13, BA14, BA24, BA25, BA32, BA44, BA45, BA46, and BA47.

Affective neuroscience is the study of how the brain processes emotions. This field combines neuroscience with the psychological study of personality, emotion, and mood. The basis of emotions and what emotions are remains an issue of debate within the field of affective neuroscience.
Frontostriatal circuits are neural pathways that connect frontal lobe regions with the basal ganglia (striatum) that mediate motor, cognitive, and behavioural functions within the brain. They receive inputs from dopaminergic, serotonergic, noradrenergic, and cholinergic cell groups that modulate information processing. Frontostriatal circuits are part of the executive functions. Executive functions include the following: selection and perception of important information, manipulation of information in working memory, planning and organization, behavioral control, adaptation to changes, and decision making. These circuits are involved in neurodegenerative disorders such as Alzheimer's disease and Parkinson's disease as well as neuropsychiatric disorders including schizophrenia, depression, obsessive compulsive disorder (OCD), and in neurodevelopmental disorder such as attention-deficit hyperactivity disorder (ADHD).

The reward system is a group of neural structures responsible for incentive salience, associative learning, and positively-valenced emotions, particularly ones involving pleasure as a core component. Reward is the attractive and motivational property of a stimulus that induces appetitive behavior, also known as approach behavior, and consummatory behavior. A rewarding stimulus has been described as "any stimulus, object, event, activity, or situation that has the potential to make us approach and consume it is by definition a reward". In operant conditioning, rewarding stimuli function as positive reinforcers; however, the converse statement also holds true: positive reinforcers are rewarding.

The ventromedial prefrontal cortex (vmPFC) is a part of the prefrontal cortex in the mammalian brain. The ventral medial prefrontal is located in the frontal lobe at the bottom of the cerebral hemispheres and is implicated in the processing of risk and fear, as it is critical in the regulation of amygdala activity in humans. It also plays a role in the inhibition of emotional responses, and in the process of decision-making and self-control. It is also involved in the cognitive evaluation of morality.
Scientific studies have found that different brain areas show altered activity in humans with major depressive disorder (MDD), and this has encouraged advocates of various theories that seek to identify a biochemical origin of the disease, as opposed to theories that emphasize psychological or situational causes. Factors spanning these causative groups include nutritional deficiencies in magnesium, vitamin D, and tryptophan with situational origin but biological impact. Several theories concerning the biologically based cause of depression have been suggested over the years, including theories revolving around monoamine neurotransmitters, neuroplasticity, neurogenesis, inflammation and the circadian rhythm. Physical illnesses, including hypothyroidism and mitochondrial disease, can also trigger depressive symptoms.
The biology of obsessive–compulsive disorder (OCD) refers biologically based theories about the mechanism of OCD. Cognitive models generally fall into the category of executive dysfunction or modulatory control. Neuroanatomically, functional and structural neuroimaging studies implicate the prefrontal cortex (PFC), basal ganglia (BG), insula, and posterior cingulate cortex (PCC). Genetic and neurochemical studies implicate glutamate and monoamine neurotransmitters, especially serotonin and dopamine.
Neuromorality is an emerging field of neuroscience that studies the connection between morality and neuronal function. Scientists use fMRI and psychological assessment together to investigate the neural basis of moral cognition and behavior. Evidence shows that the central hub of morality is the prefrontal cortex guiding activity to other nodes of the neuromoral network. A spectrum of functional characteristics within this network to give rise to both altruistic and psychopathological behavior. Evidence from the investigation of neuromorality has applications in both clinical neuropsychiatry and forensic neuropsychiatry.

Bipolar disorder is an affective disorder characterized by periods of elevated and depressed mood. The cause and mechanism of bipolar disorder is not yet known, and the study of its biological origins is ongoing. Although no single gene causes the disorder, a number of genes are linked to increase risk of the disorder, and various gene environment interactions may play a role in predisposing individuals to developing bipolar disorder. Neuroimaging and postmortem studies have found abnormalities in a variety of brain regions, and most commonly implicated regions include the ventral prefrontal cortex and amygdala. Dysfunction in emotional circuits located in these regions have been hypothesized as a mechanism for bipolar disorder. A number of lines of evidence suggests abnormalities in neurotransmission, intracellular signalling, and cellular functioning as possibly playing a role in bipolar disorder.
Social cognitive neuroscience is the scientific study of the biological processes underpinning social cognition. Specifically, it uses the tools of neuroscience to study "the mental mechanisms that create, frame, regulate, and respond to our experience of the social world". Social cognitive neuroscience uses the epistemological foundations of cognitive neuroscience, and is closely related to social neuroscience. Social cognitive neuroscience employs human neuroimaging, typically using functional magnetic resonance imaging (fMRI). Human brain stimulation techniques such as transcranial magnetic stimulation and transcranial direct-current stimulation are also used. In nonhuman animals, direct electrophysiological recordings and electrical stimulation of single cells and neuronal populations are utilized for investigating lower-level social cognitive processes.