Related Research Articles

In molecular biology, messenger ribonucleic acid (mRNA) is a single-stranded molecule of RNA that corresponds to the genetic sequence of a gene, and is read by a ribosome in the process of synthesizing a protein.
Polyadenylation is the addition of a poly(A) tail to an RNA transcript, typically a messenger RNA (mRNA). The poly(A) tail consists of multiple adenosine monophosphates; in other words, it is a stretch of RNA that has only adenine bases. In eukaryotes, polyadenylation is part of the process that produces mature mRNA for translation. In many bacteria, the poly(A) tail promotes degradation of the mRNA. It, therefore, forms part of the larger process of gene expression.
In molecular biology, the five-prime cap is a specially altered nucleotide on the 5′ end of some primary transcripts such as precursor messenger RNA. This process, known as mRNA capping, is highly regulated and vital in the creation of stable and mature messenger RNA able to undergo translation during protein synthesis. Mitochondrial mRNA and chloroplastic mRNA are not capped.
In cell biology, a granule is a small particle barely visible by light microscopy. The term is most often used to describe a secretory vesicle containing important components of cell phyisology. Examples of granules include granulocytes, platelet granules, insulin granules, germane granules, starch granules, and stress granules.

In cellular biology, stress granules are biomolecular condensates in the cytosol composed of proteins and RNAs that assemble into 0.1–2 μm membraneless organelles when the cell is under stress. The mRNA molecules found in stress granules are stalled translation pre-initiation complexes associated with 40S ribosomal subunits, translation initiation factors, poly(A)+ mRNAs and RNA-binding proteins (RBPs). While they are membraneless organelles, stress granules have been proposed to be associated with the endoplasmatic reticulum. There are also nuclear stress granules. This article is about the cytosolic variety.

The exosome complex is a multi-protein intracellular complex capable of degrading various types of RNA molecules. Exosome complexes are found in both eukaryotic cells and archaea, while in bacteria a simpler complex called the degradosome carries out similar functions.

5′-3′ exoribonuclease 1 (Xrn1) is a protein that in humans is encoded by the XRN1 gene. Xrn1 hydrolyses RNA in the 5′ to 3′ direction.

Polyadenylate-binding protein 1 is a protein that in humans is encoded by the PABPC1 gene. The protein PABP1 binds mRNA and facilitates a variety of functions such as transport into and out of the nucleus, degradation, translation, and stability. There are two separate PABP1 proteins, one which is located in the nucleus (PABPN1) and the other which is found in the cytoplasm (PABPC1). The location of PABP1 affects the role of that protein and its function with RNA.
Poly(A)-specific ribonuclease (PARN), also known as polyadenylate-specific ribonuclease or deadenylating nuclease (DAN), is an enzyme that in humans is encoded by the PARN gene.

Regulator of nonsense transcripts 2 is a protein that in humans is encoded by the UPF2 gene.

Trinucleotide repeat-containing gene 6A protein is a protein that in humans is encoded by the TNRC6A gene.

mRNA-decapping enzyme 2 is a protein that in humans is encoded by the DCP2 gene.
mRNA-decapping enzyme 1A is a protein that in humans is encoded by the DCP1A gene.

mRNA-decapping enzyme 1B is a protein that in humans is encoded by the DCP1B gene.
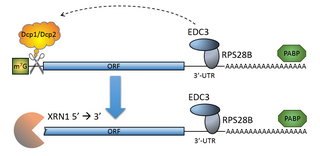
The process of messenger RNA decapping consists of hydrolysis of the 5' cap structure on the RNA exposing a 5' monophosphate. In eukaryotes, this 5' monophosphate is a substrate for the 5' exonuclease Xrn1 and the mRNA is quickly destroyed. There are many situations which may lead to the removal of the cap, some of which are discussed below.

The mRNA decapping complex is a protein complex in eukaryotic cells responsible for removal of the 5' cap. The active enzyme of the decapping complex is the bilobed Nudix family enzyme Dcp2, which hydrolyzes 5' cap and releases 7mGDP and a 5'-monophosphorylated mRNA. This decapped mRNA is inhibited for translation and will be degraded by exonucleases. The core decapping complex is conserved in eukaryotes. Dcp2 is activated by Decapping Protein 1 (Dcp1) and in higher eukaryotes joined by the scaffold protein VCS. Together with many other accessory proteins, the decapping complex assembles in P-bodies in the cytoplasm.
M7GpppX diphosphatase (EC 3.6.1.59, DcpS, m7GpppX pyrophosphatase, m7GpppN m7GMP phosphohydrolase) is an enzyme with systematic name m7G5'ppp5'N m7GMP phosphohydrolase. This enzyme catalyses the following chemical reaction
M7GpppN-mRNA hydrolase (EC 3.6.1.62, DCP2, NUDT16, D10 protein, D9 protein, D10 decapping enzyme, decapping enzyme) is an enzyme with systematic name m7GpppN-mRNA m7GDP phosphohydrolase. This enzyme catalyses the following chemical reaction
In cellular biology, a chromatoid body is a dense structure in the cytoplasm of male germ cells. It is composed mainly of RNA and RNA-binding proteins and is thus a type of RNP granule. Chromatoid body-like granules first appear in spermatocytes and condense into a single granule in round spermatids. The structure disappears again when spermatids start to elongate. The chromatoid body is crucial for spermatogenesis, but its exact role in the process is not known. Following significant strides in the understanding of small non-coding RNA mediated gene regulation and Piwi-interacting RNA (piRNA) and their roles in germline development, the function of chromatoid bodies (CBs) has been somewhat elucidated. However, due to similarities with RNP granules found in somatic cells – such as stress granules and processing bodies – chromatoid body is thought to be involved in post-transcriptional regulation of gene expression. Postmeiotic germ cell differentiation induces the accumulation of piRNAs and proteins of piRNA machinery along with several distinct RNA regulator proteins. Although evidence suggests CB involvement in mRNA regulation and small RNA mediated gene regulation, the mechanism of action remains obscure.
Roy R. Parker is a biochemist who has been an active investigator in science since the 1970s. He is currently a Distinguished Professor of Chemistry and Biochemistry and Cech-Leinwand Endowed Chair of Biochemistry at the University of Colorado Boulder. Throughout his life, Parker has contributed a vast degree of knowledge to research and studies of biochemistry. His current focus includes the biogenesis, function, and degradation of multiple forms of RNA in eukaryotes. Parker aims to use his research to understand how various diseases and pathologies result from abnormalities in RNA. In 2012, Parker was elected to the National Academy of Sciences in Biochemistry.
References
- ↑ Luo Y, Na Z, Slavoff SA (May 2018). "P-Bodies: Composition, Properties, and Functions". Biochemistry. 57 (17): 2424–2431. doi:10.1021/acs.biochem.7b01162. PMC 6296482 . PMID 29381060.
- 1 2 Kulkarni M, Ozgur S, Stoecklin G (February 2010). "On track with P-bodies". Biochemical Society Transactions. 38 (Pt 1): 242–251. doi:10.1042/BST0380242. PMID 20074068.
- ↑ Brengues M, Teixeira D, Parker R (October 2005). "Movement of eukaryotic mRNAs between polysomes and cytoplasmic processing bodies". Science. 310 (5747): 486–489. Bibcode:2005Sci...310..486B. doi:10.1126/science.1115791. PMC 1863069 . PMID 16141371.
- ↑ Bhattacharyya SN, Habermacher R, Martine U, Closs EI, Filipowicz W (June 2006). "Relief of microRNA-mediated translational repression in human cells subjected to stress". Cell. 125 (6): 1111–1124. doi: 10.1016/j.cell.2006.04.031 . PMID 16777601. S2CID 18353167.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 Hubstenberger A, Courel M, Bénard M, Souquere S, Ernoult-Lange M, Chouaib R, et al. (October 2017). "P-Body Purification Reveals the Condensation of Repressed mRNA Regulons". Molecular Cell. 68 (1): 144–157.e5. doi: 10.1016/j.molcel.2017.09.003 . PMID 28965817.
- ↑ Long, Roy M.; McNally, Mark T. (2003-05-01). "mRNA Decay: X (XRN1) Marks the Spot". Molecular Cell. 11 (5): 1126–1128. doi: 10.1016/S1097-2765(03)00198-9 . ISSN 1097-2765.
- 1 2 3 Sheth U, Parker R (May 2003). "Decapping and decay of messenger RNA occur in cytoplasmic processing bodies". Science. 300 (5620): 805–808. Bibcode:2003Sci...300..805S. doi:10.1126/science.1082320. PMC 1876714 . PMID 12730603.
- ↑ Brengues, Muriel; Teixeira, Daniela; Parker, Roy (2005-10-21). "Movement of Eukaryotic mRNAs Between Polysomes and Cytoplasmic Processing Bodies". Science. 310 (5747): 486–489. doi:10.1126/science.1115791. ISSN 0036-8075. PMC 1863069 . PMID 16141371.
- ↑ Horvathova, Ivana; Voigt, Franka; Kotrys, Anna V.; Zhan, Yinxiu; Artus-Revel, Caroline G.; Eglinger, Jan; Stadler, Michael B.; Giorgetti, Luca; Chao, Jeffrey A. (2017-11-02). "The Dynamics of mRNA Turnover Revealed by Single-Molecule Imaging in Single Cells". Molecular Cell. 68 (3): 615–625.e9. doi: 10.1016/j.molcel.2017.09.030 . ISSN 1097-2765. PMID 29056324.
- ↑ Cougot N, Bhattacharyya SN, Tapia-Arancibia L, Bordonné R, Filipowicz W, Bertrand E, Rage F (December 2008). "Dendrites of mammalian neurons contain specialized P-body-like structures that respond to neuronal activation". The Journal of Neuroscience. 28 (51): 13793–13804. doi:10.1523/JNEUROSCI.4155-08.2008. PMC 6671906 . PMID 19091970.
- 1 2 Bashkirov VI, Scherthan H, Solinger JA, Buerstedde JM, Heyer WD (February 1997). "A mouse cytoplasmic exoribonuclease (mXRN1p) with preference for G4 tetraplex substrates". The Journal of Cell Biology. 136 (4): 761–773. doi:10.1083/jcb.136.4.761. PMC 2132493 . PMID 9049243.
- ↑ Eystathioy T, Chan EK, Tenenbaum SA, Keene JD, Griffith K, Fritzler MJ (April 2002). "A phosphorylated cytoplasmic autoantigen, GW182, associates with a unique population of human mRNAs within novel cytoplasmic speckles". Molecular Biology of the Cell. 13 (4): 1338–1351. doi:10.1091/mbc.01-11-0544. PMC 102273 . PMID 11950943.
- 1 2 3 4 5 6 7 8 9 10 11 Ingelfinger D, Arndt-Jovin DJ, Lührmann R, Achsel T (December 2002). "The human LSm1-7 proteins colocalize with the mRNA-degrading enzymes Dcp1/2 and Xrnl in distinct cytoplasmic foci". RNA. 8 (12): 1489–1501. doi:10.1017/S1355838202021726. PMC 1370355 . PMID 12515382.
- 1 2 3 van Dijk E, Cougot N, Meyer S, Babajko S, Wahle E, Séraphin B (December 2002). "Human Dcp2: a catalytically active mRNA decapping enzyme located in specific cytoplasmic structures". The EMBO Journal. 21 (24): 6915–6924. doi:10.1093/emboj/cdf678. PMC 139098 . PMID 12486012.
- ↑ Gibbings DJ, Ciaudo C, Erhardt M, Voinnet O (September 2009). "Multivesicular bodies associate with components of miRNA effector complexes and modulate miRNA activity". Nature Cell Biology. 11 (9): 1143–1149. doi:10.1038/ncb1929. PMID 19684575. S2CID 205286867.
- ↑ Hoyle NP, Castelli LM, Campbell SG, Holmes LE, Ashe MP (October 2007). "Stress-dependent relocalization of translationally primed mRNPs to cytoplasmic granules that are kinetically and spatially distinct from P-bodies". The Journal of Cell Biology. 179 (1): 65–74. doi:10.1083/jcb.200707010. PMC 2064737 . PMID 17908917.
- ↑ Liu J, Valencia-Sanchez MA, Hannon GJ, Parker R (July 2005). "MicroRNA-dependent localization of targeted mRNAs to mammalian P-bodies". Nature Cell Biology. 7 (7): 719–723. doi:10.1038/ncb1274. PMC 1855297 . PMID 15937477.
- ↑ Liu J, Rivas FV, Wohlschlegel J, Yates JR, Parker R, Hannon GJ (December 2005). "A role for the P-body component GW182 in microRNA function". Nature Cell Biology. 7 (12): 1261–1266. doi:10.1038/ncb1333. PMC 1804202 . PMID 16284623.
- ↑ Sen GL, Blau HM (June 2005). "Argonaute 2/RISC resides in sites of mammalian mRNA decay known as cytoplasmic bodies". Nature Cell Biology. 7 (6): 633–636. doi:10.1038/ncb1265. PMID 15908945. S2CID 6085169.
- ↑ Eystathioy T, Jakymiw A, Chan EK, Séraphin B, Cougot N, Fritzler MJ (October 2003). "The GW182 protein colocalizes with mRNA degradation associated proteins hDcp1 and hLSm4 in cytoplasmic GW bodies". RNA. 9 (10): 1171–1173. doi:10.1261/rna.5810203. PMC 1370480 . PMID 13130130.
- ↑ Horvathova, Ivana; Voigt, Franka; Kotrys, Anna V.; Zhan, Yinxiu; Artus-Revel, Caroline G.; Eglinger, Jan; Stadler, Michael B.; Giorgetti, Luca; Chao, Jeffrey A. (2017-11-02). "The Dynamics of mRNA Turnover Revealed by Single-Molecule Imaging in Single Cells". Molecular Cell. 68 (3): 615–625.e9. doi: 10.1016/j.molcel.2017.09.030 . ISSN 1097-2765. PMID 29056324.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 Youn JY, Dunham WH, Hong SJ, Knight JD, Bashkurov M, Chen GI, et al. (February 2018). "High-Density Proximity Mapping Reveals the Subcellular Organization of mRNA-Associated Granules and Bodies". Molecular Cell. 69 (3): 517–532.e11. doi: 10.1016/j.molcel.2017.12.020 . PMID 29395067.
- 1 2 3 4 Kedersha N, Stoecklin G, Ayodele M, Yacono P, Lykke-Andersen J, Fritzler MJ, et al. (June 2005). "Stress granules and processing bodies are dynamically linked sites of mRNP remodeling". The Journal of Cell Biology. 169 (6): 871–884. doi:10.1083/jcb.200502088. PMC 2171635 . PMID 15967811.
- 1 2 Zhang B, Shi Q, Varia SN, Xing S, Klett BM, Cook LA, Herman PK (July 2016). "The Activity-Dependent Regulation of Protein Kinase Stability by the Localization to P-Bodies". Genetics. 203 (3): 1191–1202. doi:10.1534/genetics.116.187419. PMC 4937477 . PMID 27182950.
- 1 2 3 4 Cougot N, Babajko S, Séraphin B (April 2004). "Cytoplasmic foci are sites of mRNA decay in human cells". The Journal of Cell Biology. 165 (1): 31–40. doi:10.1083/jcb.200309008. PMC 2172085 . PMID 15067023.
- 1 2 3 Eystathioy T, Jakymiw A, Chan EK, Séraphin B, Cougot N, Fritzler MJ (October 2003). "The GW182 protein colocalizes with mRNA degradation associated proteins hDcp1 and hLSm4 in cytoplasmic GW bodies". RNA. 9 (10): 1171–1173. doi:10.1261/rna.5810203. PMC 1370480 . PMID 13130130.
- ↑ Eystathioy T, Chan EK, Tenenbaum SA, Keene JD, Griffith K, Fritzler MJ (April 2002). "A phosphorylated cytoplasmic autoantigen, GW182, associates with a unique population of human mRNAs within novel cytoplasmic speckles". Molecular Biology of the Cell. 13 (4): 1338–1351. doi:10.1091/mbc.01-11-0544. PMC 102273 . PMID 11950943.
- ↑ Yang Z, Jakymiw A, Wood MR, Eystathioy T, Rubin RL, Fritzler MJ, Chan EK (November 2004). "GW182 is critical for the stability of GW bodies expressed during the cell cycle and cell proliferation". Journal of Cell Science. 117 (Pt 23): 5567–5578. doi: 10.1242/jcs.01477 . PMID 15494374.
- ↑ Kucherenko MM, Shcherbata HR (January 2018). "Stress-dependent miR-980 regulation of Rbfox1/A2bp1 promotes ribonucleoprotein granule formation and cell survival". Nature Communications. 9 (1): 312. Bibcode:2018NatCo...9..312K. doi:10.1038/s41467-017-02757-w. PMC 5778076 . PMID 29358748.