
Misumena vatia is a species of crab spider with a holarctic distribution. In North America, it is called the goldenrod crab spider or flower (crab) spider, as it is commonly found hunting in goldenrod sprays and milkweed plants. They are called crab spiders because of their unique ability to walk sideways as well as forwards and backwards. Both males and females of this species progress through several molts before reaching their adult sizes, though females must molt more to reach their larger size. Females can grow up to 10 mm (0.39 in) while males are quite small, reaching 5 mm (0.20 in) at most. Misumena vatia are usually yellow or white or a pattern of these two colors. They may also present with pale green or pink instead of yellow, again, in a pattern with white. They have the ability to change between these colors based on their surroundings through the molting process. They have a complex visual system, with eight eyes, that they rely on for prey capture and for their color-changing abilities. Sometimes, if Misumena vatia consumes colored prey, the spider itself will take on that color.

Philodromus is a genus of philodromid crab spiders. Spiders in this genus are distinctively flattened.
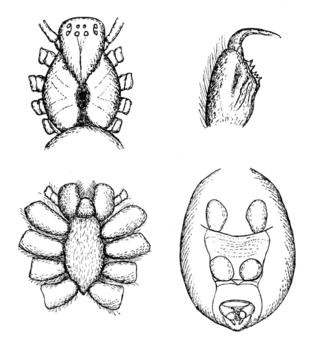
Hickmania is a monotypic genus of Australian cribellate araneomorph spiders in the family Gradungulidae, containing only the Tasmanian cave spider. The genus was first described by Willis J. Gertsch in 1958, and has been found only in Tasmania. It is the last of an old Gondwanan lineage, long since separated from its closest relatives in South America. It is an icon species for faunal conservation in Tasmania, and is named in honor of V. V. Hickman, a professor at the University of Tasmania, who specialized in spiders. The species name is derived from the Ancient Greek τρωγλοδύτης (troglodytes), meaning "cave-dweller".

The Entelegynae or entelegynes are a subgroup of araneomorph spiders, the largest of the two main groups into which the araneomorphs were traditionally divided. Females have a genital plate (epigynum) and a "flow through" fertilization system; males have complex palpal bulbs. Molecular phylogenetic studies have supported the monophyly of Entelegynae.

Sexual cannibalism is when an animal, usually the female, cannibalizes its mate prior to, during, or after copulation. It is a trait observed in many arachnid orders, several insect and crustacean clades, gastropods, and some snake species. Several hypotheses to explain this seemingly paradoxical behavior have been proposed. The adaptive foraging hypothesis, aggressive spillover hypothesis and mistaken identity hypothesis are among the proposed hypotheses to explain how sexual cannibalism evolved. This behavior is believed to have evolved as a manifestation of sexual conflict, occurring when the reproductive interests of males and females differ. In many species that exhibit sexual cannibalism, the female consumes the male upon detection. Females of cannibalistic species are generally hostile and unwilling to mate; thus many males of these species have developed adaptive behaviors to counteract female aggression.

Unicorn ("one horn", in Latin) is a genus of goblin spiders from South America, containing seven species that occur predominantly in high elevation, semi-desert regions of Bolivia, Chile, and Argentina. Individuals are relatively large for goblin spiders, measuring up to 3.0 mm (0.12 in) in body length. The genus name refers to a characteristic pointed projection between the eyes and jaws of males. In at least one species, broken-off tips of the male pedipalps have been found within the genitalia of females, postulated as a means of sperm competition. Unicorn possesses several traits that suggest it is a relatively "primitive" member of the Oonopidae, and is classified with other similar, soft-bodied goblin spiders in the subfamily Sulsulinae.

Pholcus phalangioides, commonly known as the cosmopolitan cellar spider, long-bodied cellar spider or one of various types called a daddy long-legs spider, is a spider of the family Pholcidae. It is also known as the skull spider, since its cephalothorax is said to resemble a human skull. This is the only spider species described by the Swiss entomologist Johann Kaspar Füssli, who first recorded it in 1775. Its common name of "daddy long-legs" should not be confused with a different arachnid group with the same common name, the harvestman (Opiliones), or the crane flies of the superfamily Tipuloidea.
Monogyny is a specialised mating system in which a male can only mate with one female throughout his lifetime, but the female may mate with more than one male. In this system, the males generally provide no paternal care. In many spider species that are monogynous, the males have two copulatory organs, which allows them to mate a maximum of twice throughout their lifetime. As is commonly seen in honeybees, ants and certain spider species, a male may put all his energy into a single copulation, knowing that this will lower his overall fitness. During copulation, monogynous males have adapted to cause self genital damage or even death to increase their chances of paternity.

Pisaurina mira, also known as the American nursery web spider, is a species of spider in the family Pisauridae. They are often mistaken for wolf spiders (Lycosidae) due to their physical resemblance. P. mira is distinguished by its unique eye arrangement of two rows.

Holocnemus pluchei, commonly known as the marbled cellar spider, is a species of Pholcidae, a family commonly referred to as "cellar spiders" or "daddy long-legs". This species is distributed across the North Pacific region of the United States, as well as in parts of North Africa, Europe, and the Mediterranean. It is considered a common household spider and builds its nest in attics, basements, and eaves of houses. Although some members of the species live in solitary webs, the majority join already existing webs and migrate to new webs multiple times throughout the course of their lives. A unique feature of H. pluchei is that while in many species of spiders, stridulation commonly occurs by males during sexual encounters, in H. pluchei, females also possess stridulatory organs, and both sexes engage in stridulation.

The two palpal bulbs – also known as palpal organs and genital bulbs – are the copulatory organs of a male spider. They are borne on the last segment of the pedipalps, giving the spider an appearance often described as like wearing boxing gloves. The palpal bulb does not actually produce sperm, being used only to transfer it to the female. Palpal bulbs are only fully developed in adult male spiders and are not completely visible until after the final moult. In the majority of species of spider, the bulbs have complex shapes and are important in identification.

Philodromus aureolus, the wandering crab spider, is a mainly European running crab spider of the family Philodromidae. The taxonomy of the species group named after Philodromus aureolus is in a state of flux and a number of new species have recently been recognised.

Stenaelurillus modestus is a species of jumping spider in the genus Stenaelurillus that lives in South Africa. It was first described in 2014 by Wanda Wesołowska. The species is a specialist that preys on Odontotermes termites, although its venom is also effective against other prey. The spider is medium-sized, with a brown cephalothorax between 2.0 and 2.7 mm in length and a black abdomen between 1.9 and 2.8 mm long. It is generally black in colour. The carapace has a border of white hairs and the abdomen is edged with bristles. It is distinguished from other members of the genus by the male's elongated palpal bulb and straight embolus, and the horseshoe-shaped depression in the female's epigyne.

Eris militaris, known commonly as the bronze jumper or bronze lake jumper, is a species of jumping spider, belonging to the Salticidae family. It is found in the United States and Canada within both suburban and rural areas. The male and female of this species can be differentiated from their size or by the coloration on their cephalothorax and abdomen. The females have a lighter cephalothorax a slightly darker abdomen with white spots. They are active in the autumn and winter season and can be found in sheltered areas within vegetation. They can also be found living within apple orchards, where insecticides may be present, which can potentially effect or alter their personality and behavior. Their diet consists of small insects, almost anything they can hold.
Agelenopsis pennsylvanica, commonly known as the Pennsylvania funnel-web spider or the Pennsylvania grass spider, is a species of spider in the family Agelenidae. The common name comes from the place that it was described, Pennsylvania, and the funnel shape of its web. Its closest relative is Agelenopsis potteri.

Leucauge mariana is a long-jawed orb weaver spider, native to Central America and South America. Its web building and sexual behavior have been studied extensively. Males perform several kinds of courtship behavior to induce females to copulate and to use their sperm.

Pardosa pseudoannulata, a member of a group of species referred to as wolf-spiders, is a non-web-building spider belonging to the family Lycosidae. P. pseudoannulata are wandering spiders that track and ambush prey and display sexual cannibalism. They are commonly encountered in farmlands across China and other East Asian countries. Their venom has properties that helps it function as an effective insecticide, and it is, therefore, a crucial pesticide control agent.

Larinia jeskovi is a species of the family of orb weaver spiders and a part of the genus Larinia. It is distributed throughout the Americas, Africa, Australia, Europe, and Asia and commonly found in wet climes such as marshes, bogs, and rainforests. Larinia jeskovi have yellow bodies with stripes and range from 5.13 to 8.70 millimeters in body length. They build their webs on plants with a small height above small bodies of waters or wetlands. After sunset and before sunrise are the typical times they hunt and build their web. Males usually occupy a female's web instead of making their own. The mating behavior is noteworthy as male spiders often mutilate external female genitalia to reduce sperm competition while female spiders resort to sexual cannibalism to counter such mechanisms. The males also follow an elaborate courtship ritual to attract the female. The bite of Larinia jeskovi is not known to be of harm to humans.
Zodarion wesolowskae is a species of ant spider in the genus Zodarion that lives in Morocco. The species was first described in 2020 by Souâd Benhalima and Robert Bosmans. Only the male has been described, although Benhalima and Bosmans suggest that the female could be one of the spiders described as Zodarion trianguliferum. The spider is small, typically 3.4 mm (0.13 in) long, with a plain brown to dark brown carapace and a black abdomen which has faint white stripes. The spider is a member of the mostafai group within the genus, which lack a tooth at the end of the embolus. It can be further distinguished from other members of the genus by its very long and thin tibial apophysis. The genus Zodarion is known to use ant mimicry for both defence against predators and to deceive ants to prey on them. This attribute could be used as a form of biological pest control.

Phrurolithus festivus is a spider in the family Phrurolithidae. It was originally distributed only in the Palaearctic and was introduced into Canada. The highly adaptable species inhabits a variety of both dry and wet habitats and can also be found in man-made habitats, but generally prefers open areas.

