
Orthohantavirus is a genus of single-stranded, enveloped, negative-sense RNA viruses in the family Hantaviridae within the order Bunyavirales. Members of this genus may be called orthohantaviruses or simply hantaviruses.

Bunyavirales is an order of segmented negative-strand RNA viruses with mainly tripartite genomes. Member viruses infect arthropods, plants, protozoans, and vertebrates. It is the only order in the class Ellioviricetes. The name Bunyavirales derives from Bunyamwera, where the original type species Bunyamwera orthobunyavirus was first discovered. Ellioviricetes is named in honor of late virologist Richard M. Elliott for his early work on bunyaviruses.

The term viral protein refers to both the products of the genome of a virus and any host proteins incorporated into the viral particle. Viral proteins are grouped according to their functions, and groups of viral proteins include structural proteins, nonstructural proteins, regulatory proteins, and accessory proteins. Viruses are non-living and do not have the means to reproduce on their own, instead depending on their host cell's machinery to do this. Thus, viruses do not code for most of the proteins required for their replication and the translation of their mRNA into viral proteins, but use proteins encoded by the host cell for this purpose.

Tick-borne encephalitis virus (TBEV) is a positive-strand RNA virus associated with tick-borne encephalitis in the genus Flavivirus.
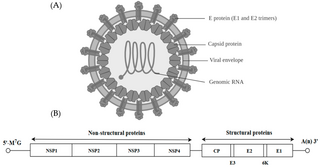
Alphavirus is a genus of RNA viruses, the sole genus in the Togaviridae family. Alphaviruses belong to group IV of the Baltimore classification of viruses, with a positive-sense, single-stranded RNA genome. There are 32 alphaviruses, which infect various vertebrates such as humans, rodents, fish, birds, and larger mammals such as horses, as well as invertebrates. Alphaviruses that could infect both vertebrates and arthropods are referred dual-host alphaviruses, while insect-specific alphaviruses such as Eilat virus and Yada yada virus are restricted to their competent arthropod vector. Transmission between species and individuals occurs mainly via mosquitoes, making the alphaviruses a member of the collection of arboviruses – or arthropod-borne viruses. Alphavirus particles are enveloped, have a 70 nm diameter, tend to be spherical, and have a 40 nm isometric nucleocapsid.

Thogotovirus is a genus of enveloped RNA viruses, one of seven genera in the virus family Orthomyxoviridae. Their single-stranded, negative-sense RNA genome has six or seven segments. Thogotoviruses are distinguished from most other orthomyxoviruses by being arboviruses – viruses that are transmitted by arthropods, in this case usually ticks. Thogotoviruses can replicate in both tick cells and vertebrate cells; one subtype has also been isolated from mosquitoes. A consequence of being transmitted by blood-sucking vectors is that the virus must spread systemically in the vertebrate host – unlike influenza viruses, which are transmitted by respiratory droplets and are usually confined to the respiratory system.

Pappataci fever is a vector-borne febrile arboviral infection caused by three serotypes of Phlebovirus. It occurs in subtropical regions of the Eastern Hemisphere. The name, pappataci fever, comes from the Italian word for sandfly; it is the union of the words pappa and taci (silent), distinguishing these insects from blood-feeding mosquitoes, which produce a typical noise while flying.

Orthonairovirus is a genus of viruses in the family Nairoviridae of the order Bunyavirales that include viruses with circular, negative-sense single stranded RNA. It got its name from the Nairobi sheep disease that affects the gastrointestinal tracts of sheep and goats. The vast majority, and perhaps all viruses in this genus are tick-borne viruses that can have human or other vertebrate hosts.

Orthobunyavirus is a genus of the Peribunyaviridae family in the order Bunyavirales. There are currently ~170 viruses recognised in this genus. These have been assembled into 103 species and 20 serogroups.
Toscana phlebovirus (TOSV) is an arbovirus belonging to Bunyavirales, an order of negative-stranded, enveloped RNA viruses. The virus can be transmitted to humans by the bite of an infected sandfly of the genus Phlebotomus. Toscana is not normally associated with disease, as indicated by high seroprevalence rates in endemic areas, but in common with other sandfly transmitted viruses such as Naples virus and Sicilian virus, infection may result in Pappataci fever, an illness with mild fever, headache and myalgia. In serious cases that go undiagnosed, acute meningitis, meningoencephalitis and encephalitis may occur. There is no specific treatment for infection, so treatment is supportive, reducing the severity of symptoms until the immune system has cleared the infection.
Dabie bandavirus, also called SFTS virus, is a tick-borne virus in the genus Bandavirus in the family Phenuiviridae, order Bunyavirales. The clinical condition it caused is known as severe fever with thrombocytopenia syndrome (SFTS). SFTS is an emerging infectious disease that was first described in northeast and central China 2009 and now has also been discovered in Japan, South Korea, Vietnam and Taiwan in 2015. SFTS has a fatality rate of 12% and as high as over 30% in some areas. The major clinical symptoms of SFTS are fever, vomiting, diarrhea, multiple organ failure, thrombocytopenia, leukopenia and elevated liver enzyme levels. Another outbreak occurred in East China in the early half of 2020.
Sangassou orthohantavirus(SANGV) is single-stranded, negative-sense RNA virus species of the genus Orthohantavirus in the Bunyavirales order. It was first isolated in an African wood mouse (Hylomyscus simus) in the forest in Guinea, West Africa in 2010. It is named for the village near where the mouse was trapped. It is the first indigenous Murinae-associated African hantavirus to be discovered.
Naples phlebovirus is an antigenic species of genus Phlebovirus within the family Phenuiviridae of the order Bunyavirales. It is an enveloped RNA virus with a tripartite genome e Uukuniemi (UUK) serogroup. The Sandfly group's natural reservoir are sandflies, while the natural reservoir for Uukuniemi is ticks. The SFNV serogroup consists of two main serocomplexes associated with disease in humans, the Naples and Sicilian serocomplexes. Sandfly fever induces myalgia, fever, and elevated liver enzymes in humans. It is difficult to diagnose outside endemic areas.
In 1954 the Hazara orthonairovirus, one of the 34 tick-borne viruses of the genus Orthonairovirus, was discovered in Pakistan in the Ixodes tick native to that region. Today this virus is studied in mice in an attempt to develop treatments for the highly pathogenic Crimean-Congo Hemorrhagic Fever virus.
The Bhanja virus is a tick-borne virus first discovered in a tick taken from a paralyzed goat in Bhanjanagar, India in 1954. Bhanja virus in humans was first documented in 1974 when Charles Calisher was working with the virus in a lab and contracted it himself. His experience with the virus was mild and included symptoms of mild aching in muscles and joints, moderate headache, slight photophobia. The Bhanja virus is a member of the Bhanja virus serocomplex and is a member of the Bunyavirales order.
Batai orthobunyavirus (BATV) is a RNA virus belonging to order Bunyavirales, genus Orthobunyavirus.
Cache Valley orthobunyavirus (CVV) is a member of the order Bunyavirales, genus Orthobunyavirus, and serogroup Bunyamwera, which was first isolated in 1956 from Culiseta inornata mosquitos collected in Utah's Cache Valley. CVV is an enveloped arbovirus, nominally 80–120 nm in diameter, whose genome is composed of three single-stranded, negative-sense RNA segments. The large segment of related bunyaviruses is approximately 6800 bases in length and encodes a probable viral polymerase. The middle CVV segment has a 4463-nucleotide sequence and the smallest segment encodes for the nucleocapsid, and a second non-structural protein. CVV has been known to cause outbreaks of spontaneous abortion and congenital malformations in ruminants such as sheep and cattle. CVV rarely infects humans, but when they are infected it has caused encephalitis and multiorgan failure.
The Punta Toro virus is a member of the genus Phlebovirus of the order Bunyavirales. It was initially isolated from patients in Colombia and two key patients in Panama. Two individual serotypes of PTV were isolated from these patients, PTV-Adames (A) and PTV-Balliet (B), with PTV-A appearing to be more virulent. PTV is considered to be relatively contained to the Americas with no cases being reported outside of this region. Along with a few other human pathogenic Phleboviruses, PTV is considered to be a significant virus in terms of public health as little information is known about its clinical effects and with further research underway, PTV could have unforeseen impacts on health and virology.
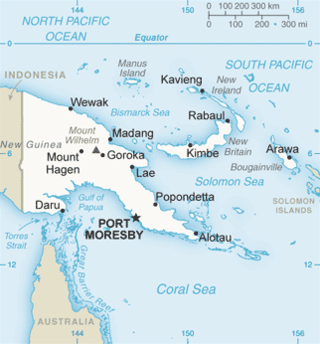
Sepik virus (SEPV) is an arthropod-borne virus (arbovirus) of the genus Flavivirus and family Flaviviridae. Flaviviridae is one of the most well characterized viral families, as it contains many well-known viruses that cause diseases that have become very prevalent in the world, like Dengue virus. The genus Flavivirus is one of the largest viral genera and encompasses over 50 viral species, including tick and mosquito borne viruses like Yellow fever virus and West Nile virus. Sepik virus is much less well known and has not been as well-classified as other viruses because it has not been known of for very long. Sepik virus was first isolated in 1966 from the mosquito Mansoniaseptempunctata, and it derives its name from the Sepik River area in Papua New Guinea, where it was first found. The geographic range of Sepik virus is limited to Papua New Guinea, due to its isolation.

Modoc virus (MODV) is a rodent-associated flavivirus. Small and enveloped, MODV contains positive single-stranded RNA. Taxonomically, MODV is part of the Flavivirus genus and Flaviviridae family. The Flavivirus genus includes nearly 80 viruses, both vector-borne and no known vector (NKV) species. Known flavivirus vector-borne viruses include Dengue virus, Yellow Fever virus, tick-borne encephalitis virus, and West Nile virus.


