
The human microbiome is the aggregate of all microbiota that reside on or within human tissues and biofluids along with the corresponding anatomical sites in which they reside, including the gastrointestinal tract, skin, mammary glands, seminal fluid, uterus, ovarian follicles, lung, saliva, oral mucosa, conjunctiva, and the biliary tract. Types of human microbiota include bacteria, archaea, fungi, protists, and viruses. Though micro-animals can also live on the human body, they are typically excluded from this definition. In the context of genomics, the term human microbiome is sometimes used to refer to the collective genomes of resident microorganisms; however, the term human metagenome has the same meaning.

An exoenzyme, or extracellular enzyme, is an enzyme that is secreted by a cell and functions outside that cell. Exoenzymes are produced by both prokaryotic and eukaryotic cells and have been shown to be a crucial component of many biological processes. Most often these enzymes are involved in the breakdown of larger macromolecules. The breakdown of these larger macromolecules is critical for allowing their constituents to pass through the cell membrane and enter into the cell. For humans and other complex organisms, this process is best characterized by the digestive system which breaks down solid food via exoenzymes. The small molecules, generated by the exoenzyme activity, enter into cells and are utilized for various cellular functions. Bacteria and fungi also produce exoenzymes to digest nutrients in their environment, and these organisms can be used to conduct laboratory assays to identify the presence and function of such exoenzymes. Some pathogenic species also use exoenzymes as virulence factors to assist in the spread of these disease-causing microorganisms. In addition to the integral roles in biological systems, different classes of microbial exoenzymes have been used by humans since pre-historic times for such diverse purposes as food production, biofuels, textile production and in the paper industry. Another important role that microbial exoenzymes serve is in the natural ecology and bioremediation of terrestrial and marine environments.

Psychrophiles or cryophiles are extremophilic organisms that are capable of growth and reproduction in low temperatures, ranging from −20 °C (−4 °F) to 20 °C (68 °F). They are found in places that are permanently cold, such as the polar regions and the deep sea. They can be contrasted with thermophiles, which are organisms that thrive at unusually high temperatures, and mesophiles at intermediate temperatures. Psychrophile is Greek for 'cold-loving', from Ancient Greek ψυχρός (psukhrós) 'cold, frozen'.

Sphingomonas was defined in 1990 as a group of Gram-negative, rod-shaped, chemoheterotrophic, strictly aerobic bacteria. They possess ubiquinone 10 as their major respiratory quinone, contain glycosphingolipids (GSLs), specifically ceramide, instead of lipopolysaccharide (LPS) in their cell envelopes, and typically produce yellow-pigmented colonies. The GSL serves to protect the bacteria from antibacterial substances. Unlike most Gram-negative bacteria, Sphingomonas cannot carry endotoxins due to the lack of lipopolysaccharides, and has a hydrophobic surface characterized by the short nature of the GSL's carbohydrate portion.

An arbuscular mycorrhiza (AM) is a type of mycorrhiza in which the symbiont fungus penetrates the cortical cells of the roots of a vascular plant forming arbuscules. Arbuscular mycorrhiza is a type of endomycorrhiza along with ericoid mycorrhiza and orchid mycorrhiza .They are characterized by the formation of unique tree-like structures, the arbuscules. In addition, globular storage structures called vesicles are often encountered.

Mycoremediation is a form of bioremediation in which fungi-based remediation methods are used to decontaminate the environment. Fungi have been proven to be a cheap, effective and environmentally sound way for removing a wide array of contaminants from damaged environments or wastewater. These contaminants include heavy metals, organic pollutants, textile dyes, leather tanning chemicals and wastewater, petroleum fuels, polycyclic aromatic hydrocarbons, pharmaceuticals and personal care products, pesticides and herbicides in land, fresh water, and marine environments.
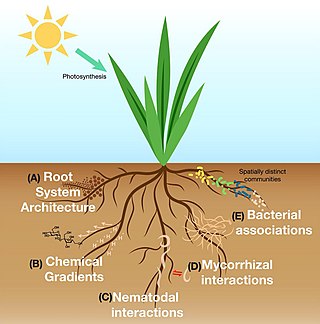
The rhizosphere is the narrow region of soil or substrate that is directly influenced by root secretions and associated soil microorganisms known as the root microbiome. Soil pores in the rhizosphere can contain many bacteria and other microorganisms that feed on sloughed-off plant cells, termed rhizodeposition, and the proteins and sugars released by roots, termed root exudates. This symbiosis leads to more complex interactions, influencing plant growth and competition for resources. Much of the nutrient cycling and disease suppression by antibiotics required by plants occurs immediately adjacent to roots due to root exudates and metabolic products of symbiotic and pathogenic communities of microorganisms. The rhizosphere also provides space to produce allelochemicals to control neighbours and relatives.
Mycolic acids are long fatty acids found in the cell walls of Mycobacteriales taxon, a group of bacteria that includes Mycobacterium tuberculosis, the causative agent of the disease tuberculosis. They form the major component of the cell wall of many Mycobacteriales species. Despite their name, mycolic acids have no biological link to fungi; the name arises from the filamentous appearance their presence gives Mycobacteriales under high magnification. The presence of mycolic acids in the cell wall also gives Mycobacteriales a distinct gross morphological trait known as "cording". Mycolic acids were first isolated by Stodola et al. in 1938 from an extract of M. tuberculosis.

Soil biology is the study of microbial and faunal activity and ecology in soil. Soil life, soil biota, soil fauna, or edaphon is a collective term that encompasses all organisms that spend a significant portion of their life cycle within a soil profile, or at the soil-litter interface. These organisms include earthworms, nematodes, protozoa, fungi, bacteria, different arthropods, as well as some reptiles, and species of burrowing mammals like gophers, moles and prairie dogs. Soil biology plays a vital role in determining many soil characteristics. The decomposition of organic matter by soil organisms has an immense influence on soil fertility, plant growth, soil structure, and carbon storage. As a relatively new science, much remains unknown about soil biology and its effect on soil ecosystems.

Surfactin is a cyclic lipopeptide, commonly used as an antibiotic for its capacity as a surfactant. It is an amphiphile capable of withstanding hydrophilic and hydrophobic environments. The Gram-positive bacterial species Bacillus subtilis produces surfactin for its antibiotic effects against competitors. Surfactin showcases antibacterial, antiviral, antifungal, and hemolytic effects.

Archaea is a domain of single-celled organisms. These microorganisms lack cell nuclei and are therefore prokaryotic. Archaea were initially classified as bacteria, receiving the name archaebacteria, but this term has fallen out of use.

Microbiota are the range of microorganisms that may be commensal, mutualistic, or pathogenic found in and on all multicellular organisms, including plants. Microbiota include bacteria, archaea, protists, fungi, and viruses, and have been found to be crucial for immunologic, hormonal, and metabolic homeostasis of their host.
Decomposition in animals is a process that begins immediately after death and involves the destruction of soft tissue, leaving behind skeletonized remains. The chemical process of decomposition is complex and involves the breakdown of soft tissue, as the body passes through the sequential stages of decomposition. Autolysis and putrefaction also play major roles in the disintegration of cells and tissues.
Soil microbiology is the study of microorganisms in soil, their functions, and how they affect soil properties. It is believed that between two and four billion years ago, the first ancient bacteria and microorganisms came about on Earth's oceans. These bacteria could fix nitrogen, in time multiplied, and as a result released oxygen into the atmosphere. This led to more advanced microorganisms, which are important because they affect soil structure and fertility. Soil microorganisms can be classified as bacteria, actinomycetes, fungi, algae and protozoa. Each of these groups has characteristics that define them and their functions in soil.
Syntrophococcus sucromutans is a Gram-negative strictly anaerobic chemoorganotrophic Bacillota. These bacteria can be found forming small chains in the habitat where it was first isolated, the rumen of cows. It is the type strain of genus Syntrophococcus and it has an uncommon one-carbon metabolic pathway, forming acetate from formate as a product of sugar oxidation.
A phytobiome consists of a plant (phyto) situated in its specific ecological area (biome), including its environment and the associated communities of organisms which inhabit it. These organisms include all macro- and micro-organisms living in, on, or around the plant including bacteria, archaea, fungi, protists, insects, animals, and other plants. The environment includes the soil, air, and climate. Examples of ecological areas are fields, rangelands, forests. Knowledge of the interactions within a phytobiome can be used to create tools for agriculture, crop management, increased health, preservation, productivity, and sustainability of cropping and forest systems.

Mycorrhiza helper bacteria (MHB) are a group of organisms that form symbiotic associations with both ectomycorrhiza and arbuscular mycorrhiza. MHBs are diverse and belong to a wide variety of bacterial phyla including both Gram-negative and Gram-positive bacteria. Some of the most common MHBs observed in studies belong to the phylas Pseudomonas and Streptomyces. MHBs have been seen to have extremely specific interactions with their fungal hosts at times, but this specificity is lost with plants. MHBs enhance mycorrhizal function, growth, nutrient uptake to the fungus and plant, improve soil conductance, aid against certain pathogens, and help promote defense mechanisms. These bacteria are naturally present in the soil, and form these complex interactions with fungi as plant root development starts to take shape. The mechanisms through which these interactions take shape are not well-understood and needs further study.

The plant microbiome, also known as the phytomicrobiome, plays roles in plant health and productivity and has received significant attention in recent years. The microbiome has been defined as "a characteristic microbial community occupying a reasonably well-defined habitat which has distinct physio-chemical properties. The term thus not only refers to the microorganisms involved but also encompasses their theatre of activity".
Hydrocarbonoclastic bacteria are a heterogeneous group of prokaryotes which can degrade and utilize hydrocarbon compounds as source of carbon and energy. Despite being present in most of environments around the world, several of these specialized bacteria live in the sea and have been isolated from polluted seawater.
An oleaginous microorganism is a type of microbe that accumulates lipid as a normal part of its metabolism. Oleaginous microbes may accumulate an array of different lipid compounds, including polyhydroxyalkanoates, triacylglycerols, and wax esters. Various microorganisms, including bacteria, fungi, and yeast, are known to accumulate lipids. These organisms are often researched for their potential use in producing fuels from waste products.



