A halophile is an extremophile that thrives in high salt concentrations. In chemical terms, halophile refers to a Lewis acidic species that has some ability to extract halides from other chemical species.

Psychrophiles or cryophiles are extremophilic organisms that are capable of growth and reproduction in low temperatures, ranging from −20 °C (−4 °F) to 20 °C (68 °F). They are found in places that are permanently cold, such as the polar regions and the deep sea. They can be contrasted with thermophiles, which are organisms that thrive at unusually high temperatures, and mesophiles at intermediate temperatures. Psychrophile is Greek for 'cold-loving', from Ancient Greek ψυχρός (psukhrós) 'cold, frozen'.

A salt lake or saline lake is a landlocked body of water that has a concentration of salts and other dissolved minerals significantly higher than most lakes. In some cases, salt lakes have a higher concentration of salt than sea water; such lakes can also be termed hypersaline lakes, and may also be pink lakes on account of their colour. An alkalic salt lake that has a high content of carbonate is sometimes termed a soda lake.

Beggiatoa is a genus of Gammaproteobacteria belonging to the order Thiotrichales, in the Pseudomonadota phylum. These bacteria form colorless filaments composed of cells that can be up to 200 µm in diameter, and are one of the largest prokaryotes on Earth. Beggiatoa are chemolithotrophic sulfur-oxidizers, using reduced sulfur species as an energy source. They live in sulfur-rich environments such as soil, both marine and freshwater, in the deep sea hydrothermal vents, and in polluted marine environments. In association with other sulfur bacteria, e.g. Thiothrix, they can form biofilms that are visible to the naked eye as mats of long white filaments; the white color is due to sulfur globules stored inside the cells.

A brine pool, sometimes called an underwater lake, deepwater or brine lake, is a volume of brine collected in a seafloor depression. The pools are dense bodies of water that have a salinity that is three to eight times greater than the surrounding ocean. Brine pools are commonly found below polar sea ice and in the deep ocean. Those below sea ice form through a process called brine rejection. For deep-sea brine pools, salt is necessary to increase the salinity gradient. The salt can come from one of two processes: the dissolution of large salt deposits through salt tectonics or geothermally heated brine issued from tectonic spreading centers.
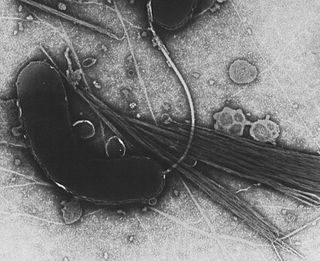
Gammaproteobacteria is a class of bacteria in the phylum Pseudomonadota. It contains about 250 genera, which makes it the most genus-rich taxon of the Prokaryotes. Several medically, ecologically, and scientifically important groups of bacteria belong to this class. All members of this class are Gram-negative. It is the most phylogenetically and physiologically diverse class of the Pseudomonadota.
Hydrogen-oxidizing bacteria are a group of facultative autotrophs that can use hydrogen as an electron donor. They can be divided into aerobes and anaerobes. The former use hydrogen as an electron donor and oxygen as an acceptor while the latter use sulphate or nitrogen dioxide as electron acceptors. Species of both types have been isolated from a variety of environments, including fresh waters, sediments, soils, activated sludge, hot springs, hydrothermal vents and percolating water.
A chemocline is a type of cline, a layer of fluid with different properties, characterized by a strong, vertical chemistry gradient within a body of water. In bodies of water where chemoclines occur, the cline separates the upper and lower layers, resulting in different properties for those layers. The lower layer shows a change in the concentration of dissolved gases and solids compared to the upper layer.
Haloplasma contractile is a halophilic, cell wall-less bacterium. It is the only known representative of a deep lineage, and is classified in its own family (Haloplasmataceae) and order (Haloplasmatales), in the class Mollicutes. In terms of genetics, the bacterium Haloplasma contractile contains a dcw gene cluster is responsible for containing all the genes of the organism and promoting peptidoglycan synthesis. Also, MreB/Mbl are specific homologous parts of this bacterium that are vital in the contractility of the cell. Regarding its physical attributes, this organism consists of a spherical body with approximately two protrusions which alternate between straight and contracted forms.

Blood Falls is an outflow of an iron oxide–tainted plume of saltwater, flowing from the tongue of Taylor Glacier onto the ice-covered surface of West Lake Bonney in the Taylor Valley of the McMurdo Dry Valleys in Victoria Land, East Antarctica.
L'Atalante basin is a hypersaline brine lake at the bottom of the Mediterranean Sea about 192 km (119 mi) west of the island of Crete. It is named for the French L'Atalante, one of the oceanographic research vessels involved in its discovery in 1993. L'Atalante and its neighbors the Urania and Discovery deep hyper saline anoxic basins (DHABs) are at most 35,000 years old. They were formed by Messinian evaporite salt deposits dissolving out of the Mediterranean Ridge and collecting in abyssal depressions about 3,000 m (9,800 ft) deep. L'Atalante is the smallest of the three; its surface begins at about 3,500 m (11,500 ft) below sea level.

A soda lake or alkaline lake is a lake on the strongly alkaline side of neutrality, typically with a pH value between 9 and 12. They are characterized by high concentrations of carbonate salts, typically sodium carbonate, giving rise to their alkalinity. In addition, many soda lakes also contain high concentrations of sodium chloride and other dissolved salts, making them saline or hypersaline lakes as well. High pH and salinity often coincide, because of how soda lakes develop. The resulting hypersaline and highly alkalic soda lakes are considered some of the most extreme aquatic environments on Earth.

The class Zetaproteobacteria is the sixth and most recently described class of the Pseudomonadota. Zetaproteobacteria can also refer to the group of organisms assigned to this class. The Zetaproteobacteria were originally represented by a single described species, Mariprofundus ferrooxydans, which is an iron-oxidizing neutrophilic chemolithoautotroph originally isolated from Kamaʻehuakanaloa Seamount in 1996 (post-eruption). Molecular cloning techniques focusing on the small subunit ribosomal RNA gene have also been used to identify a more diverse majority of the Zetaproteobacteria that have as yet been unculturable.

A brinicle is a downward-growing hollow tube of ice enclosing a plume of descending brine that is formed beneath developing sea ice.
Sulfurimonas is a bacterial genus within the class of Campylobacterota, known for reducing nitrate, oxidizing both sulfur and hydrogen, and containing Group IV hydrogenases. This genus consists of four species: Sulfurimonas autorophica, Sulfurimonas denitrificans, Sulfurimonas gotlandica, and Sulfurimonas paralvinellae. The genus' name is derived from "sulfur" in Latin and "monas" from Greek, together meaning a “sulfur-oxidizing rod”. The size of the bacteria varies between about 1.5-2.5 μm in length and 0.5-1.0 μm in width. Members of the genus Sulfurimonas are found in a variety of different environments which include deep sea-vents, marine sediments, and terrestrial habitats. Their ability to survive in extreme conditions is attributed to multiple copies of one enzyme. Phylogenetic analysis suggests that members of the genus Sulfurimonas have limited dispersal ability and its speciation was affected by geographical isolation rather than hydrothermal composition. Deep ocean currents affect the dispersal of Sulfurimonas spp., influencing its speciation. As shown in the MLSA report of deep-sea hydrothermal vents Campylobacterota, Sulfurimonas has a higher dispersal capability compared with deep sea hydrothermal vent thermophiles, indicating allopatric speciation.

Haloquadratum walsbyi is a species of the genus Haloquadratum, within the archaea domain known for its square halophilic nature. First discovered in a brine pool in the Sinai peninsula of Egypt, H. walsbyi is noted for its flat, square-shaped cells, and its unusual ability to survive in aqueous environments with high concentrations of sodium chloride and magnesium chloride. The species' genus name Haloquadratum translates from Greek and Latin as "salt square". This archaean is also commonly referred to as "Walsby's Square Bacterium" because of its identifying square shape which makes it unique. In accordance with its name, Haloquadratum walsbyi are most abundantly observed in salty environments.

Sea Ice Microbial Communities (SIMCO) refer to groups of microorganisms living within and at the interfaces of sea ice at the poles. The ice matrix they inhabit has strong vertical gradients of salinity, light, temperature and nutrients. Sea ice chemistry is most influenced by the salinity of the brine which affects the pH and the concentration of dissolved nutrients and gases. The brine formed during the melting sea ice creates pores and channels in the sea ice in which these microbes can live. As a result of these gradients and dynamic conditions, a higher abundance of microbes are found in the lower layer of the ice, although some are found in the middle and upper layers. Despite this extreme variability in environmental conditions, the taxonomical community composition tends to remain consistent throughout the year, until the ice melts.

The hydrothermal vent microbial community includes all unicellular organisms that live and reproduce in a chemically distinct area around hydrothermal vents. These include organisms in the microbial mat, free floating cells, or bacteria in an endosymbiotic relationship with animals. Chemolithoautotrophic bacteria derive nutrients and energy from the geological activity at Hydrothermal vents to fix carbon into organic forms. Viruses are also a part of the hydrothermal vent microbial community and their influence on the microbial ecology in these ecosystems is a burgeoning field of research.
An anchialine system is a landlocked body of water with a subterranean connection to the ocean. Depending on its formation, these systems can exist in one of two primary forms: pools or caves. The primary differentiating characteristics between pools and caves is the availability of light; cave systems are generally aphotic while pools are euphotic. The difference in light availability has a large influence on the biology of a given system. Anchialine systems are a feature of coastal aquifers which are density stratified, with water near the surface being fresh or brackish, and saline water intruding from the coast at depth. Depending on the site, it is sometimes possible to access the deeper saline water directly in the anchialine pool, or sometimes it may be accessible by cave diving.
A sea ice brine pocket is an area of fluid sea water with a high salt concentration trapped in sea ice as it freezes. Due to the nature of their formation, brine pockets are most commonly found in areas below −2 °C (28 °F), where it is sufficiently cold for seawater to freeze and form sea ice. Though the high salinity and low light conditions of brine pockets create a challenging environment for marine mammals, brine pockets serve as a habitat to various microbes. Sampling and studying these pockets requires specialized equipment and alterations to methodologies to accommodate the hyper-saline conditions and subzero temperatures.

