Alternative splicing, or alternative RNA splicing, or differential splicing, is an alternative splicing process during gene expression that allows a single gene to code for multiple proteins. In this process, particular exons of a gene may be included within or excluded from the final, processed messenger RNA (mRNA) produced from that gene. This means the exons are joined in different combinations, leading to different (alternative) mRNA strands. Consequently, the proteins translated from alternatively spliced mRNAs will contain differences in their amino acid sequence and, often, in their biological functions.

A spliceosome is a large ribonucleoprotein (RNP) complex found primarily within the nucleus of eukaryotic cells. The spliceosome is assembled from small nuclear RNAs (snRNA) and numerous proteins. Small nuclear RNA (snRNA) molecules bind to specific proteins to form a small nuclear ribonucleoprotein complex, which in turn combines with other snRNPs to form a large ribonucleoprotein complex called a spliceosome. The spliceosome removes introns from a transcribed pre-mRNA, a type of primary transcript. This process is generally referred to as splicing. An analogy is a film editor, who selectively cuts out irrelevant or incorrect material from the initial film and sends the cleaned-up version to the director for the final cut.
The 5′ untranslated region is the region of a messenger RNA (mRNA) that is directly upstream from the initiation codon. This region is important for the regulation of translation of a transcript by differing mechanisms in viruses, prokaryotes and eukaryotes. While called untranslated, the 5′ UTR or a portion of it is sometimes translated into a protein product. This product can then regulate the translation of the main coding sequence of the mRNA. In many organisms, however, the 5′ UTR is completely untranslated, instead forming complex secondary structure to regulate translation.

Dosage compensation is the process by which organisms equalize the expression of genes between members of different biological sexes. Across species, different sexes are often characterized by different types and numbers of sex chromosomes. In order to neutralize the large difference in gene dosage produced by differing numbers of sex chromosomes among the sexes, various evolutionary branches have acquired various methods to equalize gene expression among the sexes. Because sex chromosomes contain different numbers of genes, different species of organisms have developed different mechanisms to cope with this inequality. Replicating the actual gene is impossible; thus organisms instead equalize the expression from each gene. For example, in humans, females (XX) silence the transcription of one X chromosome of each pair, and transcribe all information from the other, expressed X chromosome. Thus, human females have the same number of expressed X-linked genes as do human males (XY), both sexes having essentially one X chromosome per cell, from which to transcribe and express genes.

DSCAM and Dscam are both abbreviations for Down syndrome cell adhesion molecule. In humans, DSCAM refers to a gene that encodes one of several protein isoforms.
RNA-binding proteins are proteins that bind to the double or single stranded RNA in cells and participate in forming ribonucleoprotein complexes. RBPs contain various structural motifs, such as RNA recognition motif (RRM), dsRNA binding domain, zinc finger and others. They are cytoplasmic and nuclear proteins. However, since most mature RNA is exported from the nucleus relatively quickly, most RBPs in the nucleus exist as complexes of protein and pre-mRNA called heterogeneous ribonucleoprotein particles (hnRNPs). RBPs have crucial roles in various cellular processes such as: cellular function, transport and localization. They especially play a major role in post-transcriptional control of RNAs, such as: splicing, polyadenylation, mRNA stabilization, mRNA localization and translation. Eukaryotic cells express diverse RBPs with unique RNA-binding activity and protein–protein interaction. According to the Eukaryotic RBP Database (EuRBPDB), there are 2961 genes encoding RBPs in humans. During evolution, the diversity of RBPs greatly increased with the increase in the number of introns. Diversity enabled eukaryotic cells to utilize RNA exons in various arrangements, giving rise to a unique RNP (ribonucleoprotein) for each RNA. Although RBPs have a crucial role in post-transcriptional regulation in gene expression, relatively few RBPs have been studied systematically.

X hyperactivation refers to the process in Drosophila by which genes on the X chromosome in male flies become twice as active as genes on the X chromosome in female flies.
Ultrabithorax (Ubx) is a homeobox gene found in insects, and is used in the regulation of patterning in morphogenesis. There are many possible products of this gene, which function as transcription factors. Ubx is used in the specification of serially homologous structures, and is used at many levels of developmental hierarchies. In Drosophila melanogaster it is expressed in the third thoracic (T3) and first abdominal (A1) segments and represses wing formation. The Ubx gene regulates the decisions regarding the number of wings and legs the adult flies will have. The developmental role of the Ubx gene is determined by the splicing of its product, which takes place after translation of the gene. The specific splice factors of a particular cell allow the specific regulation of the developmental fate of that cell, by making different splice variants of transcription factors. In D. melanogaster, at least six different isoforms of Ubx exist.
Splicing factor 1 also known as zinc finger protein 162 (ZFM162) is a protein that in humans is encoded by the SF1 gene.

HuD otherwise known as ELAV-like protein 4 is a protein that in humans is encoded by the ELAVL4 gene.

Transformer-2 protein homolog alpha is a protein that in humans is encoded by the TRA2A gene.
Splicing factor 45 is a protein that in humans is encoded by the RBM17 gene.

Notch proteins are a family of type-1 transmembrane proteins that form a core component of the Notch signaling pathway, which is highly conserved in metazoans. The Notch extracellular domain mediates interactions with DSL family ligands, allowing it to participate in juxtacrine signaling. The Notch intracellular domain acts as a transcriptional activator when in complex with CSL family transcription factors. Members of this Type 1 transmembrane protein family share several core structures, including an extracellular domain consisting of multiple epidermal growth factor (EGF)-like repeats and an intracellular domain transcriptional activation domain (TAD). Notch family members operate in a variety of different tissues and play a role in a variety of developmental processes by controlling cell fate decisions. Much of what is known about Notch function comes from studies done in Caenorhabditis elegans (C.elegans) and Drosophila melanogaster. Human homologs have also been identified, but details of Notch function and interactions with its ligands are not well known in this context.

RoX RNA is a non-coding RNA (ncRNA) present in the male-specific lethal (MSL) complex and is required for sex dosage compensation in Drosophila. As males only contain one X chromosome, male flies dosage compensate for the X chromosome by hyper-transcribing the X chromosome. This is achieved by the MSL complex binding to the X chromosome and inducing histone H4 lysine 16 acetylation and allows for the formation of euchromatin. These ncRNAs were first discovered in RNA extracted from neuronal cells.

The developmentally active and heat shock inducible hsromega or hsrω gene in Drosophila produces multiple long non-coding RNA transcripts. This gene is transcriptionally active in almost all cell types of Drosophila and is the most actively induced following heat shock. A unique feature of the hsromega gene, which led to discovery of the 93D puff in 1970, is its singular inducibility with benzamide and a variety of other amides.

In molecular biology the DM domain is a protein domain first discovered in the doublesex proteins of Drosophila melanogaster and is also seen in C. elegans and mammalian proteins. In D. melanogaster the doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides. These proteins are believed to function as transcription factors on downstream sex-determination genes, especially on neuroblast differentiation and yolk protein genes transcription.

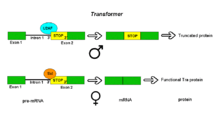
Female-specific protein transformer is a protein that in Drosophila melanogaster is encoded by the tra gene. Unlike the related tra2 protein, it is only produced in females.

Homeotic protein bicoid is encoded by the bcd maternal effect gene in Drosophilia. Homeotic protein bicoid concentration gradient patterns the anterior-posterior (A-P) axis during Drosophila embryogenesis. Bicoid was the first protein demonstrated to act as a morphogen. Although bicoid is important for the development of Drosophila and other higher dipterans, it is absent from most other insects, where its role is accomplished by other genes.
Vrille (vri) is a bZIP transcription factor found on chromosome 2 in Drosophila melanogaster. Vrille mRNA and protein product (VRI) oscillate predictably on a 24-hour timescale and interact with other circadian clock genes to regulate circadian rhythms in Drosophila. It is also a regulator in embryogenesis; it is expressed in multiple cell types during multiple stages in development, coordinating embryonic dorsal/ventral polarity, wing-vein differentiation, and ensuring tracheal integrity. It is also active in the embryonic gut but the precise function there is unknown. Mutations in vri alter circadian period and cause circadian arrhythmicity and developmental defects in Drosophila.

Transformer (tra) is a family of genes which regulate sex determination in insects such as flies. Among its effects, it regulates differences between males and females in Drosophila fruit flies.