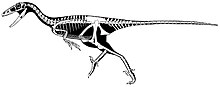
Latenivenatrix mcmasterae? (van der Reest & Currie, 2017)
Stenonychosaurus (meaning "narrow claw lizard") is a genus of troodontiddinosaur from the Late CretaceousDinosaur Park Formation of Alberta, Canada, as well as possibly the Two Medicine Formation. The type and only species, S. inequalis, was named by Charles Mortram Sternberg in 1932, based on a foot, fragments of a hand, and some caudal vertebrae from the Late Cretaceous of Alberta. S. inequalis was reassigned in 1987 by Phil Currie to the genus Troodon, which was reverted by the recognition of Stenonychosaurus as a separate genus from the possibly dubiousTroodon in 2017 by Evans et al. and also later in the same year by Van der Reest and Currie.
The first specimens currently assigned to Troodon that were not teeth were both found by Sternberg in 1928, in the Dinosaur Park Formation of Alberta. The first was named Stenonychosaurus inequalis by Sternberg in 1932, based on a foot, fragments of a hand, and some tail vertebrae. A remarkable feature of these remains was the enlarged claw on the second toe, which is now recognized as characteristic of early paravians. Sternberg initially classified Stenonychosaurus as a member of the family Coeluridae. The second, a partial lower jaw bone, was described by Gilmore (1932) as a new species of lizard which he named Polyodontosaurus grandis. Later, in 1951, Sternberg recognized P. grandis as a possible synonym of Troodon, and speculated that since Stenonychosaurus had a "very peculiar pes" and Troodon "equally unusual teeth", they may be closely related. No comparable specimens were available at that time to test the idea.
The holotype of Stenonychosaurus inequalis, CMN 8539, is a partial skeleton consisting of six caudal vertebrae, hand bones, the distal end of the left tibia and astragalus, and a complete left foot. A couple of cranial specimens were also referred to the genus: UALVP 52611 (a nearly complete skull roof) and TMP 1986.036.0457 (a partial braincase).
A more complete skeleton of Stenonychosaurus was described by Dale Russell in 1969 from the Dinosaur Park Formation, which eventually formed the scientific foundation for a famous life-sized sculpture of Stenonychosaurus accompanied by its fictional, humanoid descendant, the "dinosauroid".[1]
Stenonychosaurus became a well-known theropod in the 1980s, when the feet and braincase were described in more detail. Along with Saurornithoides, it formed the family Saurornithoididae. Based on differences in tooth structure, and the extremely fragmentary nature of the original Troodon formosus specimens, saurornithoidids were thought to be close relatives while Troodon was considered a dubious possible relative of the family. Phil Currie, reviewing the pertinent specimens in 1987, showed that supposed differences in tooth and jaw structure among troodontids and saurornithoidids were based on age and position of the tooth in the jaw, rather than a difference in species. He reclassified Stenonychosaurus inequalis as well as Polyodontosaurus grandis and Pectinodon bakkeri as junior synonyms of Troodon formosus. Currie also made Saurornithoididae a junior synonym of Troodontidae.[2] In 1988, Gregory S. Paul went farther and included Saurornithoides mongoliensis in the genus Troodon as T. mongoliensis,[3] but this reclassification, along with many other unilateral synonymizations of well known genera, was not adopted by other researchers. Currie's classification of all North American troodontid material in the single species Troodon formosus became widely adopted by other paleontologists, and all of the specimens once called Stenonychosaurus were referred to as Troodon in the scientific literature through the early 21st century.
However, the concept that all Late Cretaceous North American troodontids belong to one species began to be questioned soon after Currie's 1987 paper was published, including by Currie himself. Currie and colleagues (1990) noted that, while they believed the Judith River troodontids were all T. formosus, troodontid fossils from other formations, such as the Hell Creek Formation and Lance Formation, might belong to different species. In 1991, George Olshevsky assigned the Lance formation fossils, which had first been named Pectinodon bakkeri but later synonymized with Troodon formosus to the species Troodon bakkeri, and several other researchers (including Currie) reverted to keeping the Dinosaur Park Formation fossils separate as Troodon inequalis.[4]
In 2011, Zanno and colleagues reviewed the convoluted history of troodontid classification in Late Cretaceous North America. They followed Longrich (2008) in treating Pectinodon bakkeri as a valid genus, and noted that it is likely the numerous Late Cretaceous specimens currently assigned to Troodon formosus almost certainly represent numerous new species, but that a more thorough review of the specimens is required. Because the holotype of T. formosus is a single tooth, this may render Troodon a nomen dubium.[5]
In 2017, Evans and colleagues, building on the work of Zanno and others, confirmed the currently undiagnostic nature of the holotype of Troodon formosus and suggested that Stenonychosaurus be used for troodontid skeletal material from the Dinosaur Park Formation.[6] Later in 2017, Van der Reest and Currie found Stenonychosaurus to be a valid genus, but reassigned much of the known material to the new genus Latenivenatrix.[7]
Many Dinosaur Park troodontid specimens once referred to Troodon have recently been referred to Stenonychosaurus and Latenivenatrix. The specimens now referred to Stenonychosaurus include several frontals (UALVP 5282, TMP 1986.078.0040, TMP 1988.050.0088, TMP 1991.036.0690), a partial dentary (TMP 1982.019.0151), and the distal portion of a left metatarsal III (TMP 1998.068.0090).
In 2021, a more comprehensive re-analysis of the morphology and stratigraphic positions of known skeletal material assigned to Stenonychosaurus and Latenivenatrix determined that several characters described as diagnostic of Latenivenatrix are in fact individually variable, that both taxa overlap stratigraphically, and that Latenivenatrix mcmasterae is a junior synonym of Stenonychosaurus inequalis.[8] This leaves S. inequalis as the only valid troodontid taxon currently identified from the Dinosaur Park Formation.
A model of the hypothetical Dinosauroid, Dinosaur Museum, Dorchester
In 1982, Dale A. Russell, then curator of vertebrate fossils at the National Museum of Canada in Ottawa, conjectured a possible evolutionary path for Stenonychosaurus, if it had not perished in the Cretaceous–Paleogene extinction event, suggesting that it could have evolved into intelligent beings similar in body plan to humans. Over geologic time, Russell noted that there had been a steady increase in the encephalization quotient or EQ (the relative brain weight when compared to other species with the same body weight) among the dinosaurs. Russell had discovered the first Troodontid skull, and noted that, while its EQ was low compared to humans, it was six times higher than that of other dinosaurs. Russell suggested that if the trend in Stenonychosaurus evolution had continued to the present, its brain case could by now measure 1,100cm3 (67cuin), comparable to that of a human (on average, 1,260cm3 (77cuin) for men and 1,130cm3 (69cuin) for women).[1]
Troodontids had semi-manipulative fingers, able to grasp and hold objects to a certain degree, and binocular vision.[1] Russell proposed that his "Dinosauroid", like members of the troodontid family, would have had large eyes and three fingers on each hand, one of which would have been partially opposed. Russell also speculated that the "Dinosauroid" would have had a toothless beak. As with most modern reptiles (and birds), he conceived of its genitalia as internal. Russell speculated that it would have required a navel, as a placenta aids the development of a large brain case. However, it would not have possessed mammary glands, and would have fed its young, as some birds do, on regurgitated food. He speculated that its language would have sounded somewhat like bird song.[1][9]
However, Russell's thought experiment has been met with criticism from other paleontologists since the 1980s, many of whom point out that his Dinosauroid is overly anthropomorphic. Gregory S. Paul (1988) and Thomas R. Holtz, Jr., consider it "suspiciously human" and Darren Naish has argued that a large-brained, highly intelligent troodontid would retain a more standard theropod body plan, with a horizontal posture and long tail, and would probably manipulate objects with the snout and feet in the manner of a bird, rather than with human-like "hands".[9]
Description
Size compared to a human
Stenonychosaurus was a small dinosaur, up to 2.5 metres (8.2ft) in length and 35 kilograms (77lb) in body mass.[10] The largest specimens are comparable in size to Deinonychus and Unenlagia.[11] They had very long, slender hind limbs, suggesting that these animals were able to run quickly. They had large, retractable, sickle-shaped claws on the second toes, which were raised off the ground when running.
Their eyes were very large (perhaps suggesting a partially nocturnal lifestyle), and slightly forward facing, giving Stenonychosaurus some degree of depth perception.[12]
Brain and inner ear
Restoration
Stenonychosaurus had one of the largest known brains of any dinosaur, relative to its body mass (comparable to modern birds).[13] This has been calculated as a cerebrum-to-brain-volume ratio 31.5% to 63% of the way from a non-avian reptile proportion to a truly avian one.[14] Additionally, it had bony cristae supporting their tympanic membranes, that were ossified at least in their top and bottom regions. The rest of the cristae were either cartilaginous or too delicate to be preserved. The metotic strut of Stenonychosaurus was enlarged from side-to-side, similar to Dromaeosaurus and primitive birds like Archaeopteryx and Hesperornis.[14]
Paleobiology
Restoration of two individuals playing in snow
Stenonychosaurus are thought to have been predators, a view supported by its sickle claw on the foot and apparently good binocular vision.
Stenonychosaurus teeth, however, are different from most other theropods. One comparative study of the feeding apparatus suggests that Stenonychosaurus could have been an omnivore.[15] The jaws met in a broad, U-shaped symphysis similar to that of an iguana, a lizard species adapted to a plant-eating lifestyle. Additionally, the teeth of Stenonychosaurus bore large serrations, each of which is called a denticle. There are pits at the intersections of the denticles, and the points of the denticles point towards the tip, or apex, of each tooth. The teeth show wear facets on their sides. Holtz (1998) also noted that characteristics used to support a predatory habit for Stenonychosaurus – the grasping hands, large brain, and stereoscopic vision – are all characteristics shared with herbivorous or omnivorousprimates and omnivorous Procyon (raccoon).
Age determination studies performed on the Two Medicine troodont using growth ring counts suggest that this dinosaur reached its adult size probably in 3–5 years.[16]
A partial skeleton has been discovered with preserved puncture marks, possibly inflicted by a predator.[17]
Dinosaur eggs and nests were discovered by John R. Horner in 1983 in the Two Medicine Formation of Montana. Varriccho et al. (2002) have described eight of these nests found to date. These are all in the collection of the Museum of the Rockies and their accession numbers are MOR 246, 299, 393, 675, 676, 750, 963, 1139. Horner (1984) found isolated bones and partial skeletons of the hypsilophodontOrodromeus very near the nests in the same horizon and described the eggs as those of Orodromeus.[18] Horner and Weishampel (1996) reexamined the embryos preserved in the eggs and determined that they were those of Troodon, not Orodromeus.[19] Varricchio et al. (1997) made this determination with even more certainty when they described a partial skeleton of an adult Troodon (MOR 748) in contact with a clutch of at least five eggs (MOR 750), probably in a brooding position.[20] Van der Reest and Currie considered it possible that the Two Medicine troodont was the same species as Stenonychosaurus.[7]
Varricchio et al. (1997) described the exact structure of the nests. They were built from sediments, they were dish shaped, about 100cm (39in) in internal diameter, and with a pronounced raised rim encircling the eggs. The more complete nests had between 16 (minimum number in MOR 246) and 24 (MOR 963) eggs. The eggs are shaped like elongated teardrops, with the more tapered ends pointed downwards and embedded about halfway in the sediment. The eggs are pitched at an angle so that, on average, the upper half is closer to the center of the nest. There is no evidence that plant matter was present in the nest.
Varricchio et al.(1997) were able to extract enough evidence from the nests to infer several characteristics of troodont reproductive biology. The results are that they appear to have had a type of reproduction that is intermediate between crocodiles and birds, as phylogeny would predict. The eggs are statistically grouped in pairs, which suggests that the animal had two functional oviducts, like crocodiles, rather than one, as in birds. Crocodiles lay many eggs that are small proportional to adult body size. Birds lay fewer, larger, eggs. The Two Medicine troodont was intermediate, laying an egg of about 0.5kg (1.1lb) for a 50kg (110lb) adult. This is 10 times larger than reptiles of the same mass, but two troodont eggs are roughly equivalent to the 1.1kg (2.4lb) egg predicted for a 50kg (110lb) bird.
Varricchio et al. also found evidence for iterative laying, where the adult might lay a pair of eggs every one or two days, and then ensured simultaneous hatching by delaying brooding until all eggs were laid. MOR 363 was found with 22 empty (hatched) eggs, and the embryos found in the eggs of MOR 246 were in very similar states of development, implying that all of the young hatched approximately simultaneously. The embryos had an advanced degree of skeletal development and empty eggs were relatively uncrushed, implying that hatchlings were precocial. The authors estimated 45 to 65 total days of adult nest attendance for laying, brooding, and hatching. [21]
Varricchio et al. (2008) examined the bone histology of Two Medicine troodont specimen MOR 748 and found that it lacked the bone resorption patterns that would indicate it was an egg-laying female. They also measured the ratio of the total volume of eggs in clutches to the body mass of the adult. They graphed correlations between this ratio and the type of parenting strategies used by extant birds and crocodiles and found that the ratio in the troodont was consistent with that in birds where only the adult male broods the eggs. From this they concluded that troodont females likely did not brood eggs, that the males did, and this may be a character shared between maniraptoran dinosaurs and basal birds.[22] However, a later analysis of avian clutch mass found that the type of parental care cannot be determined using conventional allometric methods such as the one used by Varricchio et al.[23]
Troodontidae is a clade of bird-like theropod dinosaurs. During most of the 20th century, troodontid fossils were few and incomplete and they have therefore been allied, at various times, with many dinosaurian lineages. More recent fossil discoveries of complete and articulated specimens, have helped to increase understanding about this group. Anatomical studies, particularly studies of the most primitive troodontids, like Sinovenator, demonstrate striking anatomical similarities with Archaeopteryx and primitive dromaeosaurids, and demonstrate that they are relatives comprising a clade called Paraves.
Troodon is a former wastebasket taxon and a potentially dubious genus of relatively small, bird-like theropod dinosaurs definitively known from the Campanian age of the Late Cretaceous period. It includes at least one species, Troodon formosus, known from Montana. Discovered in October 1855, T. formosus was among the first dinosaurs found in North America, although it was thought to be a lizard until 1877. Several well-known troodontid specimens from the Dinosaur Park Formation in Alberta were once believed to be members of this genus. However, recent analyses in 2017 have found this genus to be undiagnostic and referred some of these specimens to the genus Stenonychosaurus some to the genus Latenivenatrix, and some to the genus Pectinodon. The genus name is Ancient Greek for "wounding tooth", referring to the teeth, which were different from those of most other theropods known at the time of their discovery. The teeth bear prominent, apically oriented serrations. These "wounding" serrations, however, are morphometrically more similar to those of herbivorous reptiles, and suggest a possibly omnivorous diet.
Saurornithoides is a genus of troodontid maniraptoran dinosaur, which lived during the Late Cretaceous period. These creatures were predators, which could run fast on their hind legs and had excellent sight and hearing. The name is derived from the Greek stems saur~ (lizard), ornith~ (bird) and eides (form), referring to its bird-like skull.
Byronosaurus is a genus of troodontid dinosaur from the Late Cretaceous Period of Mongolia.
Philip John Currie is a Canadian palaeontologist and museum curator who helped found the Royal Tyrrell Museum of Palaeontology in Drumheller, Alberta and is now a professor at the University of Alberta in Edmonton. In the 1980s, he became the director of the Canada-China Dinosaur Project, the first cooperative palaeontological partnering between China and the West since the Central Asiatic Expeditions in the 1920s, and helped describe some of the first feathered dinosaurs. He is one of the primary editors of the influential Encyclopedia of Dinosaurs, and his areas of expertise include theropods, the origin of birds, and dinosaurian migration patterns and herding behavior. He was one of the models for palaeontologist Alan Grant in the film Jurassic Park.
Orodromeus is a genus of herbivorous orodromine thescelosaurid dinosaur from the Late Cretaceous of North America. Only one species is known, the type species Orodromeus makelai.
Hagryphus is a monospecific genus of caenagnathid dinosaur from southern Utah that lived during the Late Cretaceous in what is now the Kaiparowits Formation of the Grand Staircase–Escalante National Monument. The type and only species, Hagryphus giganteus, is known only from an incomplete but articulated left manus and the distal portion of the left radius. It was named in 2005 by Lindsay E. Zanno and Scott D. Sampson. Hagryphus has an estimated length of 2.4–3 metres and weight of 50 kilograms.
Pectinodon is a genus of troodontid theropod dinosaurs from the end of the Maastrichtian age of the Late Cretaceous period (66 mya). It currently contains a single valid species, Pectinodon bakkeri, known only from teeth.
Polyodontosaurus is a potentially dubious genus of troodontid dinosaur named in 1932 by Charles W. Gilmore for a left dentary from the Dinosaur Park Formation. It had been considered a synonym of Stenonychosaurus or Troodon for a significant time, before being declared a nomen dubium. The only known species is the type, P. grandis.
Citipati is a genus of oviraptorid dinosaur that lived in Asia during the Late Cretaceous period, about 75 million to 71 million years ago. It is mainly known from the Ukhaa Tolgod locality at the Djadochta Formation, where the first remains were collected during the 1990s. The genus and type species Citipati osmolskae were named and described in 2001. A second species from the adjacent Zamyn Khondt locality may also exist. Citipati is one of the best-known oviraptorids thanks to a number of well-preserved specimens, including individuals found in brooding positions atop nests of eggs, though most of them were initially referred to the related Oviraptor. These nesting specimens have helped to solidify the link between non-avian dinosaurs and birds.
Troodontinae is a subfamily of troodontid dinosaurs. The subfamily was first used in 2017 for the group of troodontids descended from the last common ancestor of Gobivenator mongoliensis and Zanabazar junior, but has been redefined to be the least inclusive clade containing Saurornithoides mongoliensis and Troodon formosus, utilizing the type species of the clade.
Continuoolithus is an oogenus of dinosaur egg found in the late Cretaceous of North America. It is most commonly known from the late Campanian of Alberta and Montana, but specimens have also been found dating to the older Santonian and the younger Maastrichtian. It was laid by an unknown type of theropod. These small eggs are similar to the eggs of oviraptorid dinosaurs, but have a distinctive type of ornamentation.
Talos is an extinct genus of carnivorous bird-like theropod dinosaur, an advanced troodontid which lived during the late Cretaceous period in the geographic area that is now Utah, United States.
Jinfengopteryginae is a subfamily of bird-like theropod dinosaurs known from the Cretaceous of Eurasia. This group includes relatively few genera, with members discovered in 2005 but the name erected in 2012. Like other troodontids, this group of dinosaurs resided in the Paraves potentially close to the Avialae.
Oohkotokia is a genus of ankylosaurid dinosaur within the subfamily Ankylosaurinae. It is known from the upper levels of the Two Medicine Formation of Montana, United States. The discovery of Oohkotokia supports that Ankylosaurine dinosaurs existed and flourished continuously in Montana and/or Alberta throughout the late Campanian and early Maastrichtian stages in the Late Cretaceous period. It was a large, heavily built, quadrupedal, herbivore, that could grow up to 5 metres (16 ft) long and weigh up to 2 metric tons.
This timeline of troodontid research is a chronological listing of events in the history of paleontology focused on the troodontids, a group of bird-like theropod dinosaurs including animals like Troodon. Troodontid remains were among the first dinosaur fossils to be reported from North America after paleontologists began performing research on the continent, specifically the genus Troodon itself. Since the type specimen of this genus was only a tooth and Troodon teeth are unusually similar to those of the unrelated thick-headed pachycephalosaurs, Troodon and its relatives would be embroiled in taxonomic confusion for over a century. Troodon was finally recognized as distinct from the pachycephalosaurs by Phil Currie in 1987. By that time many other species now recognized as troodontid had been discovered but had been classified in the family Saurornithoididae. Since these families were the same but the Troodontidae named first, it carries scientific legitimacy.
Elongatoolithidae is an oofamily of fossil eggs, representing the eggs of oviraptorosaurs. They are known for their highly elongated shape. Elongatoolithids have been found in Europe, Asia, and both North and South America.
Latenivenatrix, meaning "hiding huntress", is a genus of large troodontid known from a single species, L. mcmasterae. Along with the contemporary Stenonychosaurus, it is known from non-tooth fossils that were formerly assigned to the now potentially dubious genus Troodon. Although described as separate, it has been considered a junior synonym of Stenonychosaurus.
The dinosauroid is a hypothetical species created by Dale A. Russell in 1982. Russell theorized that if a dinosaur such as Stenonychosaurus had not perished in the Cretaceous–Paleogene extinction event, its descendants might have evolved to fill the same ecological niche as humans. While the theory has been met with criticism from other scientists, the dinosauroid has been featured widely in books and documentaries since the theory's inception.
↑ Currie, P. (2005). "Theropods, including birds." in Currie and Koppelhus (eds). Dinosaur Provincial Park, a spectacular ecosystem revealed, Part Two, Flora and Fauna from the park. Indiana University Press, Bloomington. Pp 367–397.
1 2 van der Reest, A. J.; Currie, P. J. (2017). "Troodontids (Theropoda) from the Dinosaur Park Formation, Alberta, with a description of a unique new taxon: implications for deinonychosaur diversity in North America". Canadian Journal of Earth Sciences. 54 (9): 919–935. Bibcode:2017CaJES..54..919V. doi:10.1139/cjes-2017-0031. hdl:1807/78296.
↑ Palmer, D., ed. (1999). The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London, UK: Marshall Editions. pp.112–113. ISBN978-1-84028-152-1.
1 2 Larsson, H.C.E. (2001). "Endocranial anatomy of Carcharodontosaurus saharicus (Theropoda: Allosauroidea) and its implications for theropod brain evolution". In Tanke, D.H.; Carpenter, K.; Skrepnick, M.W. (eds.). Mesozoic Vertebrate Life. Indiana University Press. pp.19–33.
↑ Holtz, Thomas R., Brinkman, Daniel L., Chandler, Chistine L. (1998) Denticle Morphometrics and a Possibly Omnivorous Feeding Habit for the Theropod Dinosaur Troodon. Gaia number 15. December 1998. pp. 159–166.
↑ Varricchio, D. J. (1993). Bone microstructure of the Upper Cretaceous theropod dinosaur Troodon formosus. J. Vertebr. Paleontol.13, 99–104. JSTOR4523488
↑ Jacobsen, A. R. 2001. Tooth-marked small theropod bone: An extremely rare trace. p. 58-63. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
↑ Horner, John R. (1984) "The nesting behavior of dinosaurs". "Scientific American", 250:130–137.
↑ Horner, John R., Weishampel, David B. (1996) "A comparative embryological study of two ornithischian dinosaurs – a correction." "Nature" 383:256–257.
↑ Varricchio, D. J.; Jackson, F.; Borkowski, J.J.; Horner, J.R. (1997). "Nest and egg clutches of the dinosaur Troodon formosus and the evolution of avian reproductive traits". Nature. 385 (6613): 247–250. Bibcode:1997Natur.385..247V. doi:10.1038/385247a0. S2CID4313286.
↑ Varricchio, David J.; Horner, John J.; Jackson, Frankie D. (2002). "Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus". Journal of Vertebrate Paleontology. 22 (3): 564–576. doi:10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2. S2CID85728452.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.