Related Research Articles

Chloroplasts are organelles that conduct photosynthesis, where the photosynthetic pigment chlorophyll captures the energy from sunlight, converts it, and stores it in the energy-storage molecules ATP and NADPH while freeing oxygen from water in plant and algal cells. They then use the ATP and NADPH to make organic molecules from carbon dioxide in a process known as the Calvin cycle. Chloroplasts carry out a number of other functions, including fatty acid synthesis, much amino acid synthesis, and the immune response in plants. The number of chloroplasts per cell varies from one, in unicellular algae, up to 100 in plants like Arabidopsis and wheat.

Plant cells are eukaryotic cells present in green plants, photosynthetic eukaryotes of the kingdom Plantae. Their distinctive features include primary cell walls containing cellulose, hemicelluloses and pectin, the presence of plastids with the capability to perform photosynthesis and store starch, a large vacuole that regulates turgor pressure, the absence of flagella or centrioles, except in the gametes, and a unique method of cell division involving the formation of a cell plate or phragmoplast that separates the new daughter cells.

A pseudopod or pseudopodium is a temporary arm-like projection of a eukaryotic cell membrane that is developed in the direction of movement. Filled with cytoplasm, pseudopodia primarily consist of actin filaments and may also contain microtubules and intermediate filaments. Pseudopods are used for motility and ingestion. They are often found in amoebas.

Symbiogenesis, endosymbiotic theory, or serial endosymbiotic theory, is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms. The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes taken one inside the other in endosymbiosis. The idea that chloroplasts were originally independent organisms that merged into a symbiotic relationship with other one-celled organisms dates to the 19th century, espoused by researchers such as Andreas Schimper.
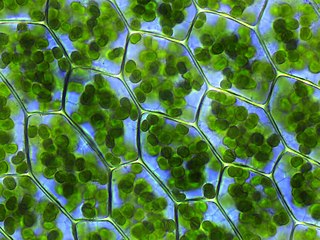
The plastid is a membrane-bound organelle found in the cells of plants, algae, and some other eukaryotic organisms. They are considered to be intracellular endosymbiotic Cyanobacteria. Examples include chloroplasts, chromoplasts, and leucoplasts.

The cytoskeleton is a complex, dynamic network of interlinking protein filaments present in the cytoplasm of all cells, including bacteria and archaea. It extends from the cell nucleus to the cell membrane and is composed of similar proteins in the various organisms. In eukaryotes, it is composed of three main components, microfilaments, intermediate filaments and microtubules, and these are all capable of rapid growth or disassembly dependent on the cell's requirements.

Microfilaments, also called actin filaments, are protein filaments in the cytoplasm of eukaryotic cells that form part of the cytoskeleton. They are primarily composed of polymers of actin, but are modified by and interact with numerous other proteins in the cell. Microfilaments are usually about 7 nm in diameter and made up of two strands of actin. Microfilament functions include cytokinesis, amoeboid movement, cell motility, changes in cell shape, endocytosis and exocytosis, cell contractility, and mechanical stability. Microfilaments are flexible and relatively strong, resisting buckling by multi-piconewton compressive forces and filament fracture by nanonewton tensile forces. In inducing cell motility, one end of the actin filament elongates while the other end contracts, presumably by myosin II molecular motors. Additionally, they function as part of actomyosin-driven contractile molecular motors, wherein the thin filaments serve as tensile platforms for myosin's ATP-dependent pulling action in muscle contraction and pseudopod advancement. Microfilaments have a tough, flexible framework which helps the cell in movement.

In cell biology, the cleavage furrow is the indentation of the cell's surface that begins the progression of cleavage, by which animal and some algal cells undergo cytokinesis, the final splitting of the membrane, in the process of cell division. The same proteins responsible for muscle contraction, actin and myosin, begin the process of forming the cleavage furrow, creating an actomyosin ring. Other cytoskeletal proteins and actin binding proteins are involved in the procedure.

Actin is a family of globular multi-functional proteins that form microfilaments in the cytoskeleton, and the thin filaments in muscle fibrils. It is found in essentially all eukaryotic cells, where it may be present at a concentration of over 100 μM; its mass is roughly 42-kDa, with a diameter of 4 to 7 nm.
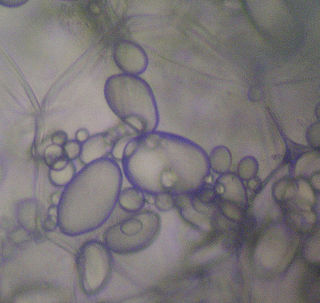
Leucoplasts are a category of plastid and as such are organelles found in plant cells. They are non-pigmented, in contrast to other plastids such as the chloroplast.
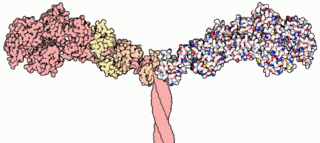
Myosins are a superfamily of motor proteins best known for their roles in muscle contraction and in a wide range of other motility processes in eukaryotes. They are ATP-dependent and responsible for actin-based motility. The term was originally used to describe a group of similar ATPases found in the cells of both striated muscle tissue and smooth muscle tissue. Following the discovery by Pollard and Korn (1973) of enzymes with myosin-like function in Acanthamoeba castellanii, a global range of divergent myosin genes have been discovered throughout the realm of eukaryotes.

Chromoplasts are plastids, heterogeneous organelles responsible for pigment synthesis and storage in specific photosynthetic eukaryotes. It is thought that like all other plastids including chloroplasts and leucoplasts they are descended from symbiotic prokaryotes.

Elaioplasts are one of the three possible forms of leucoplasts, sometimes broadly referred to as such. The main function of elaioplasts are synthesis and storage of fatty acids, terpenes, and other lipids, and they can be found in the embryonic leaves of oilseeds, citrus fruits, as well as the anthers of many flowering plants.

Cytoplasmic streaming, also called protoplasmic streaming and cyclosis, is the flow of the cytoplasm inside the cell, driven by forces from the cytoskeleton. It is likely that its function is, at least in part, to speed up the transport of molecules and organelles around the cell. It is usually observed in large plant and animal cells, greater than approximately 0.1 mm. In smaller cells, the diffusion of molecules is more rapid, but diffusion slows as the size of the cell increases, so larger cells may need cytoplasmic streaming for efficient function.
Heteroplasmy is the presence of more than one type of organellar genome within a cell or individual. It is an important factor in considering the severity of mitochondrial diseases. Because most eukaryotic cells contain many hundreds of mitochondria with hundreds of copies of mitochondrial DNA, it is common for mutations to affect only some mitochondria, leaving most unaffected.

Plasmodesmata are microscopic channels which traverse the cell walls of plant cells and some algal cells, enabling transport and communication between them. Plasmodesmata evolved independently in several lineages, and species that have these structures include members of the Charophyceae, Charales, Coleochaetales and Phaeophyceae, as well as all embryophytes, better known as land plants. Unlike animal cells, almost every plant cell is surrounded by a polysaccharide cell wall. Neighbouring plant cells are therefore separated by a pair of cell walls and the intervening middle lamella, forming an extracellular domain known as the apoplast. Although cell walls are permeable to small soluble proteins and other solutes, plasmodesmata enable direct, regulated, symplastic transport of substances between cells. There are two forms of plasmodesmata: primary plasmodesmata, which are formed during cell division, and secondary plasmodesmata, which can form between mature cells.

Motor proteins are a class of molecular motors that can move along the cytoplasm of animal cells. They convert chemical energy into mechanical work by the hydrolysis of ATP. Flagellar rotation, however, is powered by a proton pump.

A tunneling nanotube(TNT) or membrane nanotube is a term that has been applied to protrusions that extend from the plasma membrane which enable different animal cells to touch over long distances, sometimes over 100 μm between T cells. Two types of structures have been called nanotubes. The first type are less than 0.7 micrometers in diameter, contain actin and carry portions of plasma membrane between cells in both directions. The second type are larger, contain both actin and microtubules, and can carry components of the cytoplasm such as vesicles and organelles between cells, including whole mitochondria. The diameter of TNTs ranges from 50 to 200 nm and they can reach lengths of several cell diameters. These structures may be involved in cell-to-cell communication, transfer of nucleic acids between cells in a tissue, and the spread of pathogens or toxins such as HIV and prions. TNTs have observed lifetimes ranging from a few minutes up to several hours, and several proteins have been implicated in their formation or inhibition.

Omegasome is a cell compartment consisting of lipid bilayer membranes enriched for phosphatidylinositol 3-phosphate and related to a process of autophagy. It is a subdomain of the endoplasmic reticulum (ER) membrane and has a morphology resembling Greek capital letter omega (Ω). Omegasomes are the sites from which phagophores form. Phagophores are sack-like structures that mature into autophagosomes that fuse with lysosomes in order to degrade the contents of the autophagosomes. The formation of omegasomes is increased as a response to starvation.

Maureen Hanson is an American molecular biologist and Liberty Hyde Bailey Professor in the Department of Molecular Biology and Genetics at Cornell University in Ithaca, New York. She is a joint member of the Section of Plant Biology and Director of the Center for Enervating Neuroimmune Disease. Her research concerns gene expression in chloroplasts and mitochondria, photosynthesis, and the molecular basis of the disease Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS).
References
- ↑ Die Gestalts- und Lageveränderung der Pflanzen-Chromatophoren, Gustav Senn, 1908, Leipzig: Wilhelm Engelmann Verlag.
- ↑ "Die Chlorophyllkörper der Selaginellen", Gottlieb Haberlandt, Flora71 (1888), pp. 291-308.
- ↑ §2, "Stromules and the dynamic nature of plastid morphology", E. Y. Kwok and M. R. Hanson, Journal of Microscopy214, #2 (May 2004), pp. 124-137, doi : 10.1111/j.0022-2720.2004.01317.x.
- ↑ Section "The Discovery and Rediscovery of Stromules" in "Stromules: Mobile Protrusions and Interconnections Between Plastids", J. C. Gray, J. A. Sullivan, J. M. Hibberd and M. R. Hansen, Plant Biology, 3, #3 (May 2001), pp. 223-233, doi : 10.1055/s-2001-15204.
- 1 2 Köhler RH, Cao J, Zipfel WR, Webb WW, Hanson MR (1997). "Exchange of protein molecules through connections between higher plant plastids". Science. 276 (5321): 2039–42. doi:10.1126/science.276.5321.2039. PMID 9197266.
- 1 2 3 Waters, Mark; Rupert G. Fray; Kevin A. Pyke (9 July 2004). "Stromule formation is dependent upon plastid size, plastid differentiation status and the density of plastids within the cell". The Plant Journal. 39 (4): 655–67. doi: 10.1111/j.1365-313X.2004.02164.x . PMID 15272881. Archived from the original on 17 December 2012. Retrieved 7 February 2010.
- ↑ Kwok EY, Hanson MR (February 2004). "In vivo analysis of interactions between GFP-labeled microfilaments and plastid stromules". BMC Plant Biol. 4: 2. doi:10.1186/1471-2229-4-2. PMC 356911 . PMID 15018639.
- ↑ Amir Sattarzadeh, Johanna Krahmer, Arnaud D. Germain and Maureen R. Hanson (2009). "A Myosin XI Tail Domain Homologous to the Yeast Myosin Vacuole-Binding Domain Interacts with Plastids and Stromules in Nicotiana benthamiana". Molecular Plant. 2 (6): 1351–8. doi: 10.1093/mp/ssp094 . PMID 19995734.CS1 maint: multiple names: authors list (link)
- ↑ Gray JC, Sullivan JA, Hibberd JM, Hansen MR (2001). "Stromules: mobile protrusions and interconnections between plastids". Plant Biology. 3 (3): 223–233. doi:10.1055/s-2001-15204.
- ↑ Köhler RH, Hanson MR (2000). "Plastid tubules of higher plants are tissue-specific and developmentally regulated". Journal of Cell Science. 113: 81–9. PMID 10591627.