Related Research Articles

Sauria is the clade containing the most recent common ancestor of archosaurs and lepidosaurs, and all its descendants. Since most molecular phylogenies recover turtles as more closely related to archosaurs than to lepidosaurs as part of Archelosauria, Sauria can be considered the crown group of diapsids, or reptiles in general. Depending on the systematics, Sauria includes all modern reptiles or most of them as well as various extinct groups.

Dibamidae or blind skinks is a family of lizards characterized by their elongated cylindrical body and an apparent lack of limbs. Female dibamids are entirely limbless and the males retain small flap-like hind limbs, which they use to grip their partner during mating. They have a rigidly fused skull, lack pterygoid teeth and external ears. Their eyes are greatly reduced, and covered with a scale.

The quadrate bone is a skull bone in most tetrapods, including amphibians, sauropsids, and early synapsids.
The quadratojugal is a skull bone present in many vertebrates, including some living reptiles and amphibians.

Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.

Adelospondyli is an order of elongated, presumably aquatic, Carboniferous amphibians. They have a robust skull roofed with solid bone, and orbits located towards the front of the skull. The limbs were almost certainly absent, although some historical sources reported them to be present. Despite the likely absence of limbs, adelospondyls retained a large part of the bony shoulder girdle. Adelospondyls have been assigned to a variety of groups in the past. They have traditionally been seen as members of the subclass Lepospondyli, related to other unusual early tetrapods such as "microsaurs", "nectrideans", and aïstopods. Analyses such as Ruta & Coates (2007) have offered an alternate classification scheme, arguing that adelospondyls were actually far removed from other lepospondyls, instead being stem-tetrapod stegocephalians closely related to the family Colosteidae.

Temnospondyli is a diverse order of small to giant tetrapods—often considered primitive amphibians—that flourished worldwide during the Carboniferous, Permian, and Triassic periods. A few species continued into the Jurassic and Cretaceous periods. Fossils have been found on every continent. During about 210 million years of evolutionary history, they adapted to a wide range of habitats, including freshwater, terrestrial, and even coastal marine environments. Their life history is well understood, with fossils known from the larval stage, metamorphosis, and maturity. Most temnospondyls were semiaquatic, although some were almost fully terrestrial, returning to the water only to breed. These temnospondyls were some of the first vertebrates fully adapted to life on land. Although temnospondyls are considered amphibians, many had characteristics, such as scales and armour-like bony plates, that distinguish them from modern amphibians (lissamphibians).

Proganochelys is an extinct, primitive stem-turtle that has been hypothesized to be the sister taxon to all other turtles creating a monophyletic group, the Casichelydia. Proganochelys was named by Georg Baur in 1887 as the oldest turtle in existence at the time. The name Proganochelys comes from the Greek word ganos meaning 'brightness', combined with prefix pro, 'before', and Greek base chelys meaning 'turtle'. Proganochelys is believed to have been around 1 meter in size and herbivorous in nature. Proganochelys was known as the most primitive stem-turtle for over a century, until the novel discovery of Odontochelys in 2008. Odontochelys and Proganochelys share unique primitive features that are not found in Casichelydia, such as teeth on the pterygoid and vomer and a plate-like coracoid.

A parietal eye, also known as a third eye or pineal eye, is a part of the epithalamus present in some vertebrates. The eye is located at the top of the head, is photoreceptive and is associated with the pineal gland, regulating circadian rhythmicity and hormone production for thermoregulation. The hole in the head which contains the eye is known as a pineal foramen or parietal foramen, since it is often enclosed by the parietal bones.

Westlothiana is a genus of reptile-like tetrapod that lived about 338 million years ago during the latest part of the Visean age of the Carboniferous. Members of the genus bore a superficial resemblance to modern-day lizards. The genus is known from a single species, Westlothiana lizziae. The type specimen was discovered in the East Kirkton Limestone at the East Kirkton Quarry, West Lothian, Scotland in 1984. This specimen was nicknamed "Lizzie the lizard" by fossil hunter Stan Wood, and this name was quickly adopted by other paleontologists and the press. When the specimen was formally named in 1990, it was given the specific name "lizziae" in homage to this nickname. However, despite its similar body shape, Westlothiana is not considered a true lizard. Westlothiana's anatomy contained a mixture of both "labyrinthodont" and reptilian features, and was originally regarded as the oldest known reptile or amniote. However, updated studies have shown that this identification is not entirely accurate. Instead of being one of the first amniotes, Westlothiana was rather a close relative of Amniota. As a result, most paleontologists since the original description place the genus within the group Reptiliomorpha, among other amniote relatives such as diadectomorphs and seymouriamorphs. Later analyses usually place the genus as the earliest diverging member of Lepospondyli, a collection of unusual tetrapods which may be close to amniotes or lissamphibians, or potentially both at the same time.

Parareptilia is a subclass or clade of basal sauropsids (reptiles), typically considered the sister taxon to Eureptilia. Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles, the first reptiles with advanced hearing systems, and the first large herbivorous reptiles. The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.
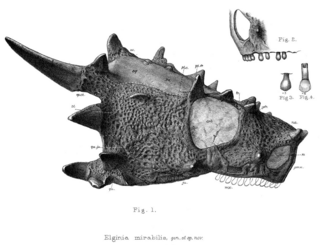
Elginia is an extinct genus of pareiasaurid known from the Late Permian of Scotland and China. It was named for the area around Elgin in Scotland, which has yielded many fossils referred to as the Elgin Reptiles.

Diplocaulus is an extinct genus of lepospondyl amphibians which lived from the Late Carboniferous to the Late Permian of North America and Africa. Diplocaulus is by far the largest and best-known of the lepospondyls, characterized by a distinctive boomerang-shaped skull. Remains attributed to Diplocaulus have been found from the Late Permian of Morocco and represent the youngest-known occurrence of a lepospondyl.

Saharastega is an extinct genus of basal temnospondyl which lived during the Late Permian period, around 251 to 260 million years ago. Remains of Saharastega, discovered by paleontologist Christian A. Sidor at the Moradi Formation in Niger, were described briefly in 2005 and more comprehensively in 2006. The description is based on a skull lacking the lower jaws.

Acherontiscus is an extinct genus of stegocephalians that lived in the Early Carboniferous of Scotland. The type and only species is Acherontiscus caledoniae, named by paleontologist Robert Carroll in 1969. Members of this genus have an unusual combination of features which makes their placement within amphibian-grade tetrapods uncertain. They possess multi-bone vertebrae similar to those of embolomeres, but also a skull similar to lepospondyls. The only known specimen of Acherontiscus possessed an elongated body similar to that of a snake or eel. No limbs were preserved, and evidence for their presence in close relatives of Acherontiscus is dubious at best. Phylogenetic analyses created by Marcello Ruta and other paleontologists in the 2000s indicate that Acherontiscus is part of Adelospondyli, closely related to other snake-like animals such as Adelogyrinus and Dolichopareias. Adelospondyls are traditionally placed within the group Lepospondyli due to their fused vertebrae. Some analyses published since 2007 have argued that adelospondyls such as Acherontiscus may not actually be lepospondyls, instead being close relatives or members of the family Colosteidae. This would indicate that they evolved prior to the split between the tetrapod lineage that leads to reptiles (Reptiliomorpha) and the one that leads to modern amphibians (Batrachomorpha). Members of this genus were probably aquatic animals that were able to swim using snake-like movements.

Rhineceps is an extinct genus of temnospondyl amphibian in the family Rhinesuchidae. Rhineceps was found in Northern Malawi in Southern Africa known only from its type species R. nyasaensis. Rhineceps was a late Permian semi-aquatic carnivore that lived in streams, rivers, lakes or lagoons. Rhineceps is an early divergent Stereopondyl within the family Rhinesuchidae, which only existed in the late Permian (Lopingian) and failed to survive the Permian-Triassic extinction unlike other stereospondyl families.

Ossinodus is an extinct genus of stem tetrapod. Fossils have been found from the Ducabrook Formation in Queensland, Australia dating back to the middle Visean stage of the Early Carboniferous (Mississippian). It was originally placed within the family Whatcheeriidae, but the absence of an intertemporal bone as suggested by a recent reconstruction of the skull based on fragmentary material may prove it to be stemward of all whatcheeriids.
Cranial kinesis is the term for significant movement of skull bones relative to each other in addition to movement at the joint between the upper and lower jaw. It is usually taken to mean relative movement between the upper jaw and the braincase.
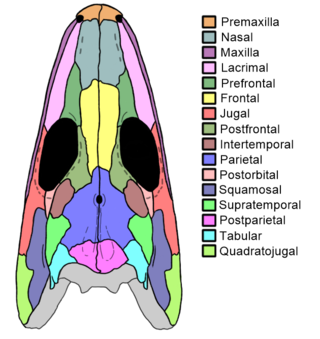
The Intertemporal bone is a paired cranial bone present in bony fish and certain extinct amphibian-grade tetrapods. It lies in the rear part of the skull, behind the eyes.

Postparietals are cranial bones present in fish and many tetrapods. Although initially a pair of bones, many lineages possess postparietals which were fused into a single bone. The postparietals were dermal bones situated along the midline of the skull, behind the parietal bones. They formed part of the rear edge of the skull roof, and the lateral edge of each postparietal often contacts the tabular and supratemporal bones. In fish, the postparietals are elongated, typically the largest components of the skull roof. Tetrapods possessed shorter postparietals, which were reduced further and shifted towards the braincase in amniotes. At several points in synapsid evolution, the postparietals fused to each other and the tabulars during embryological development. This fusion produces the interparietal bone, which is inherited by mammals. Postparietals are common in extinct amphibians and early reptiles. However, most living amphibians and living reptiles lack postparietal bones, with a few exceptions.
References
- ↑ Qiao, Tuo; Zhu, Min (2010). "Cranial morphology of the Silurian sarcopterygian Guiyu oneiros (Gnathostomata: Osteichthyes)". Science China Earth Sciences. 53 (12): 1836–1848. doi:10.1007/s11430-010-4089-6. ISSN 1674-7313. S2CID 54963054.
- ↑ Parrington, F.R. (1937). "V.— A note on the supratemporal and tabular bones in reptiles". Annals and Magazine of Natural History. 20 (115): 69–76. doi:10.1080/00222933708655315. ISSN 0374-5481.
- ↑ Schoch, Rainer R. (2014). "Amphibian skull evolution: The developmental and functional context of simplification, bone loss and heterotopy". Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 322 (8): 619–630. doi:10.1002/jez.b.22599.
- ↑ Gaffney, Eugene S.; Meeker, Lorraine J. (1983-03-01). "Skull morphology of the oldest turtles: a preliminary description of Proganochelys quenstedti". Journal of Vertebrate Paleontology. 3 (1): 25–28. doi:10.1080/02724634.1983.10011953. ISSN 0272-4634.
- ↑ Bever, G. S.; Lyson, Tyler R.; Field, Daniel J.; Bhullar, Bhart-Anjan S. (2015). "Evolutionary origin of the turtle skull". Nature. 525 (7568): 239–242. doi:10.1038/nature14900. ISSN 1476-4687. S2CID 4401555.
| | This anatomy article is a stub. You can help Wikipedia by expanding it. |