
A pheromone is a secreted or excreted chemical factor that triggers a social response in members of the same species. Pheromones are chemicals capable of acting like hormones outside the body of the secreting individual, to affect the behavior of the receiving individuals. There are alarm pheromones, food trail pheromones, sex pheromones, and many others that affect behavior or physiology. Pheromones are used by many organisms, from basic unicellular prokaryotes to complex multicellular eukaryotes. Their use among insects has been particularly well documented. In addition, some vertebrates, plants and ciliates communicate by using pheromones. The ecological functions and evolution of pheromones are a major topic of research in the field of chemical ecology.

The olfactory nerve, also known as the first cranial nerve, cranial nerve I, or simply CN I, is a cranial nerve that contains sensory nerve fibers relating to the sense of smell.
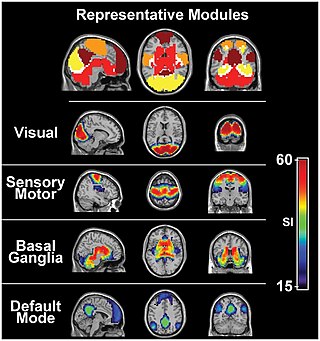
The sensory nervous system is a part of the nervous system responsible for processing sensory information. A sensory system consists of sensory neurons, neural pathways, and parts of the brain involved in sensory perception and interoception. Commonly recognized sensory systems are those for vision, hearing, touch, taste, smell, balance and visceral sensation. Sense organs are transducers that convert data from the outer physical world to the realm of the mind where people interpret the information, creating their perception of the world around them.

The flehmen response, also called the flehmen position, flehmen reaction, flehmen grimace, flehming, or flehmening, is a behavior in which an animal curls back its upper lip exposing its front teeth, inhales with the nostrils usually closed, and then often holds this position for several seconds. It may be performed over a sight or substance of particular interest to the animal, or may be performed with the neck stretched and the head held high in the air.

The olfactory bulb is a neural structure of the vertebrate forebrain involved in olfaction, the sense of smell. It sends olfactory information to be further processed in the amygdala, the orbitofrontal cortex (OFC) and the hippocampus where it plays a role in emotion, memory and learning. The bulb is divided into two distinct structures: the main olfactory bulb and the accessory olfactory bulb. The main olfactory bulb connects to the amygdala via the piriform cortex of the primary olfactory cortex and directly projects from the main olfactory bulb to specific amygdala areas. The accessory olfactory bulb resides on the dorsal-posterior region of the main olfactory bulb and forms a parallel pathway. Destruction of the olfactory bulb results in ipsilateral anosmia, while irritative lesions of the uncus can result in olfactory and gustatory hallucinations.
A chemoreceptor, also known as chemosensor, is a specialized sensory receptor which transduces a chemical substance to generate a biological signal. This signal may be in the form of an action potential, if the chemoreceptor is a neuron, or in the form of a neurotransmitter that can activate a nerve fiber if the chemoreceptor is a specialized cell, such as taste receptors, or an internal peripheral chemoreceptor, such as the carotid bodies. In physiology, a chemoreceptor detects changes in the normal environment, such as an increase in blood levels of carbon dioxide (hypercapnia) or a decrease in blood levels of oxygen (hypoxia), and transmits that information to the central nervous system which engages body responses to restore homeostasis.

The olfactory system or sense of smell is the sensory system used for smelling (olfaction). Olfaction is one of the special senses, that have directly associated specific organs. Most mammals and reptiles have a main olfactory system and an accessory olfactory system. The main olfactory system detects airborne substances, while the accessory system senses fluid-phase stimuli.

An olfactory receptor neuron (ORN), also called an olfactory sensory neuron (OSN), is a sensory neuron within the olfactory system.

The olfactory epithelium is a specialized epithelial tissue inside the nasal cavity that is involved in smell. In humans, it measures 5 cm2 (0.78 sq in) and lies on the roof of the nasal cavity about 7 cm (2.8 in) above and behind the nostrils. The olfactory epithelium is the part of the olfactory system directly responsible for detecting odors.

The glomerulus is a spherical structure located in the olfactory bulb of the brain where synapses form between the terminals of the olfactory nerve and the dendrites of mitral, periglomerular and tufted cells. Each glomerulus is surrounded by a heterogeneous population of juxtaglomerular neurons and glial cells.

The rhinarium is the furless skin surface surrounding the external openings of the nostrils in many mammals. Commonly it is referred to as the tip of the snout, and breeders of cats and dogs sometimes use the term nose leather. Informally, it may be called a "truffle", "wet snout," or "wet nose” because its surface is moist in some species: for example, healthy dogs and cats.

Vomeronasal type-1 receptor 1 is a protein that in humans is encoded by the VN1R1 gene.

The sense of smell, or olfaction, is the special sense through which smells are perceived. The sense of smell has many functions, including detecting desirable foods, hazards, and pheromones, and plays a role in taste.
Olfactory memory refers to the recollection of odors. Studies have found various characteristics of common memories of odor memory including persistence and high resistance to interference. Explicit memory is typically the form focused on in the studies of olfactory memory, though implicit forms of memory certainly supply distinct contributions to the understanding of odors and memories of them. Research has demonstrated that the changes to the olfactory bulb and main olfactory system following birth are extremely important and influential for maternal behavior. Mammalian olfactory cues play an important role in the coordination of the mother infant bond, and the following normal development of the offspring. Maternal breast odors are individually distinctive, and provide a basis for recognition of the mother by her offspring.
A sense is a biological system used by an organism for sensation, the process of gathering information about the surroundings through the detection of stimuli. Although, in some cultures, five human senses were traditionally identified as such, many more are now recognized. Senses used by non-human organisms are even greater in variety and number. During sensation, sense organs collect various stimuli for transduction, meaning transformation into a form that can be understood by the brain. Sensation and perception are fundamental to nearly every aspect of an organism's cognition, behavior and thought.

Vomeronasal receptors are a class of olfactory receptors that putatively function as receptors for pheromones. Pheromones have evolved in all animal phyla, to signal sex and dominance status, and are responsible for stereotypical social and sexual behaviour among members of the same species. In mammals, these chemical signals are believed to be detected primarily by the vomeronasal organ (VNO), a chemosensory organ located at the base of the nasal septum.
Odor molecules are detected by the olfactory receptors in the olfactory epithelium of the nasal cavity. Each receptor type is expressed within a subset of neurons, from which they directly connect to the olfactory bulb in the brain. Olfaction is essential for survival in most vertebrates; however, the degree to which an animal depends on smell is highly varied. Great variation exists in the number of OR genes among vertebrate species, as shown through bioinformatic analyses. This diversity exists by virtue of the wide-ranging environments that they inhabit. For instance, dolphins that are secondarily adapted to an aquatic niche possess a considerably smaller subset of genes than most mammals. OR gene repertoires have also evolved in relation to other senses, as higher primates with well-developed vision systems tend to have a smaller number of OR genes. As such, investigating the evolutionary changes of OR genes can provide useful information on how genomes respond to environmental changes. Differences in smell sensitivity are also dependent on the anatomy of the olfactory apparatus, such as the size of the olfactory bulb and epithelium.
Pherines, also known as vomeropherines, are odorless synthetic neuroactive steroids that engage nasal chemosensory receptors and induce dose-dependent and reversible pharmacological and behavioral effects. Pherines target human chemosensory receptors and possibly other receptors such as the GABAA receptor and influence central nervous system activity.

Insect olfaction refers to the function of chemical receptors that enable insects to detect and identify volatile compounds for foraging, predator avoidance, finding mating partners and locating oviposition habitats. Thus, it is the most important sensation for insects. Most important insect behaviors must be timed perfectly which is dependent on what they smell and when they smell it. For example, olfaction is essential for locating host plants and hunting prey in many species of insects, such as the moth Deilephila elpenor and the wasp Polybia sericea, respectively.
In zoology, copulation is animal sexual behavior in which a male introduces sperm into the female's body, especially directly into her reproductive tract. This is an aspect of mating. Many animals that live in water use external fertilization, whereas internal fertilization may have developed from a need to maintain gametes in a liquid medium in the Late Ordovician epoch. Internal fertilization with many vertebrates occurs via cloacal copulation, known as cloacal kiss, while most mammals copulate vaginally, and many basal vertebrates reproduce sexually with external fertilization.



