
Plankton are the diverse collection of organisms found in water that are unable to propel themselves against a current. The individual organisms constituting plankton are called plankters. In the ocean, they provide a crucial source of food to many small and large aquatic organisms, such as bivalves, fish, and baleen whales.
Zooplankton are the animal component of the planktonic community, having to consume other organisms to thrive. Plankton are aquatic organisms that are unable to swim effectively against currents. Consequently, they drift or are carried along by currents in the ocean, or by currents in seas, lakes or rivers.
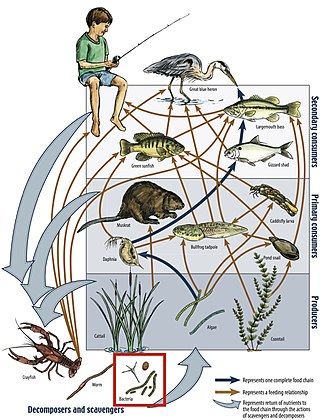
A food web is the natural interconnection of food chains and a graphical representation of what-eats-what in an ecological community. Ecologists can broadly define all life forms as either autotrophs or heterotrophs, based on their trophic levels, the position that they occupy in the food web. To maintain their bodies, grow, develop, and to reproduce, autotrophs produce organic matter from inorganic substances, including both minerals and gases such as carbon dioxide. These chemical reactions require energy, which mainly comes from the Sun and largely by photosynthesis, although a very small amount comes from bioelectrogenesis in wetlands, and mineral electron donors in hydrothermal vents and hot springs. These trophic levels are not binary, but form a gradient that includes complete autotrophs, which obtain their sole source of carbon from the atmosphere, mixotrophs, which are autotrophic organisms that partially obtain organic matter from sources other than the atmosphere, and complete heterotrophs that must feed to obtain organic matter.

Isotope analysis is the identification of isotopic signature, abundance of certain stable isotopes of chemical elements within organic and inorganic compounds. Isotopic analysis can be used to understand the flow of energy through a food web, to reconstruct past environmental and climatic conditions, to investigate human and animal diets, for food authentification, and a variety of other physical, geological, palaeontological and chemical processes. Stable isotope ratios are measured using mass spectrometry, which separates the different isotopes of an element on the basis of their mass-to-charge ratio.

Energy flow is the flow of energy through living things within an ecosystem. All living organisms can be organized into producers and consumers, and those producers and consumers can further be organized into a food chain. Each of the levels within the food chain is a trophic level. In order to more efficiently show the quantity of organisms at each trophic level, these food chains are then organized into trophic pyramids. The arrows in the food chain show that the energy flow is unidirectional, with the head of an arrow indicating the direction of energy flow; energy is lost as heat at each step along the way.

Bioturbation is defined as the reworking of soils and sediments by animals or plants. It includes burrowing, ingestion, and defecation of sediment grains. Bioturbating activities have a profound effect on the environment and are thought to be a primary driver of biodiversity. The formal study of bioturbation began in the 1800s by Charles Darwin experimenting in his garden. The disruption of aquatic sediments and terrestrial soils through bioturbating activities provides significant ecosystem services. These include the alteration of nutrients in aquatic sediment and overlying water, shelter to other species in the form of burrows in terrestrial and water ecosystems, and soil production on land.

The soil food web is the community of organisms living all or part of their lives in the soil. It describes a complex living system in the soil and how it interacts with the environment, plants, and animals.

A riparian zone or riparian area is the interface between land and a river or stream. In some regions, the terms riparian woodland, riparian forest, riparian buffer zone,riparian corridor, and riparian strip are used to characterize a riparian zone. The word riparian is derived from Latin ripa, meaning "river bank".

River ecosystems are flowing waters that drain the landscape, and include the biotic (living) interactions amongst plants, animals and micro-organisms, as well as abiotic (nonliving) physical and chemical interactions of its many parts. River ecosystems are part of larger watershed networks or catchments, where smaller headwater streams drain into mid-size streams, which progressively drain into larger river networks. The major zones in river ecosystems are determined by the river bed's gradient or by the velocity of the current. Faster moving turbulent water typically contains greater concentrations of dissolved oxygen, which supports greater biodiversity than the slow-moving water of pools. These distinctions form the basis for the division of rivers into upland and lowland rivers.

Ecological stoichiometry considers how the balance of energy and elements influences living systems. Similar to chemical stoichiometry, ecological stoichiometry is founded on constraints of mass balance as they apply to organisms and their interactions in ecosystems. Specifically, how does the balance of energy and elements affect and how is this balance affected by organisms and their interactions. Concepts of ecological stoichiometry have a long history in ecology with early references to the constraints of mass balance made by Liebig, Lotka, and Redfield. These earlier concepts have been extended to explicitly link the elemental physiology of organisms to their food web interactions and ecosystem function.

Paleolimnology is a scientific sub-discipline closely related to both limnology and paleoecology. Paleolimnological studies focus on reconstructing the past environments of inland waters using the geologic record, especially with regard to events such as climatic change, eutrophication, acidification, and internal ontogenic processes.
Trophic cascades are powerful indirect interactions that can control entire ecosystems, occurring when a trophic level in a food web is suppressed. For example, a top-down cascade will occur if predators are effective enough in predation to reduce the abundance, or alter the behavior of their prey, thereby releasing the next lower trophic level from predation.

Cross-boundary subsidies are caused by organisms or materials that cross or traverse habitat patch boundaries, subsidizing the resident populations. The transferred organisms and materials may provide additional predators, prey, or nutrients to resident species, which can affect community and food web structure. Cross-boundary subsidies of materials and organisms occur in landscapes composed of different habitat patch types, and so depend on characteristics of those patches and on the boundaries in between them. Human alteration of the landscape, primarily through fragmentation, has the potential to alter important cross-boundary subsidies to increasingly isolated habitat patches. Understanding how processes that occur outside of habitat patches can affect populations within them may be important to habitat management.

The wrack zone or wrack line is a coastal feature where organic material and other debris are deposited at high tide. This zone acts as a natural input of marine resources into a terrestrial system, providing food and habitat for a variety of coastal organisms.

The viral shunt is a mechanism that prevents marine microbial particulate organic matter (POM) from migrating up trophic levels by recycling them into dissolved organic matter (DOM), which can be readily taken up by microorganisms. The DOM recycled by the viral shunt pathway is comparable to the amount generated by the other main sources of marine DOM.

Compared to terrestrial environments, marine environments have biomass pyramids which are inverted at the base. In particular, the biomass of consumers is larger than the biomass of primary producers. This happens because the ocean's primary producers are tiny phytoplankton which grow and reproduce rapidly, so a small mass can have a fast rate of primary production. In contrast, many significant terrestrial primary producers, such as mature forests, grow and reproduce slowly, so a much larger mass is needed to achieve the same rate of primary production.

Benthic-pelagic coupling are processes that connect the benthic zone and the pelagic zone through the exchange of energy, mass, or nutrients. These processes play a prominent role in both freshwater and marine ecosystems and are influenced by a number of chemical, biological, and physical forces that are crucial to functions from nutrient cycling to energy transfer in food webs.

Aquatic macroinvertebrates are insects in their nymph and larval stages, snails, worms, crayfish, and clams that spend at least part of their lives in water. These insects play a large role in freshwater ecosystems by recycling nutrients as well as providing food to higher trophic levels.
The Hynes Award for New Investigators is awarded by the Society for Freshwater Science and recognizes an excellent academic research paper in the freshwater sciences by a scientist less than five years after their terminal graduate degree. Recipients of the award have gone on to become leading senior researchers, serving as science advisors to various governments and states, and held leadership positions in national and international scientific societies.

David M. Post is a research scientist and academic administrator. He is currently a professor of Ecology and Evolutionary Biology at Yale University and the Vice President ., Dean of Faculty, and Visiting Wong Ngit Liong Professor at Yale-NUS College, the first liberal arts college in Singapore. Post is an aquatic ecologist who studies food webs, evolution, and stable isotopes in lakes and rivers in Connecticut and Kenya.


