Ecology is the natural science of the relationships among living organisms, including humans, and their physical environment. Ecology considers organisms at the individual, population, community, ecosystem, and biosphere levels. Ecology overlaps with the closely related sciences of biogeography, evolutionary biology, genetics, ethology, and natural history.

Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.

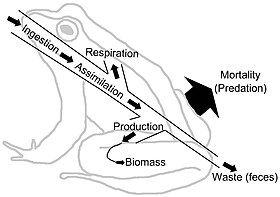
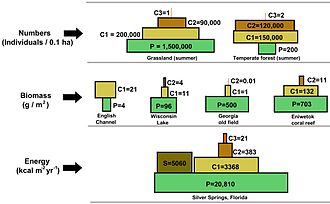
Biomass is the mass of living biological organisms in a given area or ecosystem at a given time. Biomass can refer to species biomass, which is the mass of one or more species, or to community biomass, which is the mass of all species in the community. It can include microorganisms, plants or animals. The mass can be expressed as the average mass per unit area, or as the total mass in the community.

In ecology, a biological interaction is the effect that a pair of organisms living together in a community have on each other. They can be either of the same species, or of different species. These effects may be short-term, or long-term, both often strongly influence the adaptation and evolution of the species involved. Biological interactions range from mutualism, beneficial to both partners, to competition, harmful to both partners. Interactions can be direct when physical contact is established or indirect, through intermediaries such as shared resources, territories, ecological services, metabolic waste, toxins or growth inhibitors. This type of relationship can be shown by net effect based on individual effects on both organisms arising out of relationship.
This glossary of ecology is a list of definitions of terms and concepts in ecology and related fields. For more specific definitions from other glossaries related to ecology, see Glossary of biology, Glossary of evolutionary biology, and Glossary of environmental science.

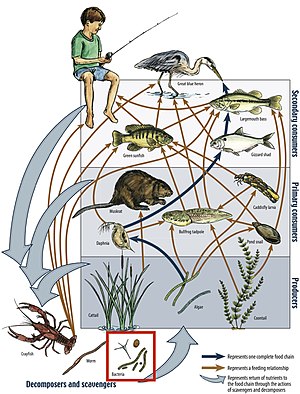
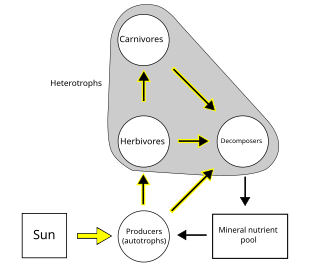
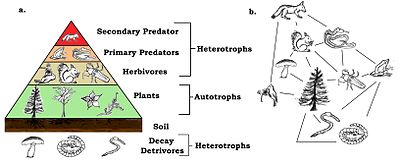
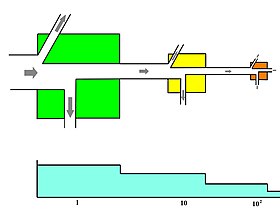
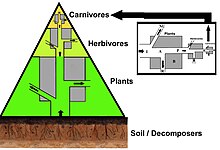
Energy flow is the flow of energy through living things within an ecosystem. All living organisms can be organized into producers and consumers, and those producers and consumers can further be organized into a food chain. Each of the levels within the food chain is a trophic level. In order to more efficiently show the quantity of organisms at each trophic level, these food chains are then organized into trophic pyramids. The arrows in the food chain show that the energy flow is unidirectional, with the head of an arrow indicating the direction of energy flow; energy is lost as heat at each step along the way.

Detritivores are heterotrophs that obtain nutrients by consuming detritus. There are many kinds of invertebrates, vertebrates, and plants that carry out coprophagy. By doing so, all these detritivores contribute to decomposition and the nutrient cycles. Detritivores should be distinguished from other decomposers, such as many species of bacteria, fungi and protists, which are unable to ingest discrete lumps of matter. Instead, these other decomposers live by absorbing and metabolizing on a molecular scale. The terms detritivore and decomposer are often used interchangeably, but they describe different organisms. Detritivores are usually arthropods and help in the process of remineralization. Detritivores perform the first stage of remineralization, by fragmenting the dead plant matter, allowing decomposers to perform the second stage of remineralization.

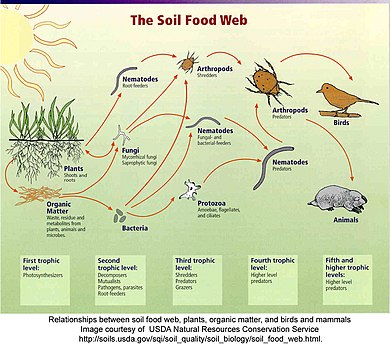
The soil food web is the community of organisms living all or part of their lives in the soil. It describes a complex living system in the soil and how it interacts with the environment, plants, and animals.

River ecosystems are flowing waters that drain the landscape, and include the biotic (living) interactions amongst plants, animals and micro-organisms, as well as abiotic (nonliving) physical and chemical interactions of its many parts. River ecosystems are part of larger watershed networks or catchments, where smaller headwater streams drain into mid-size streams, which progressively drain into larger river networks. The major zones in river ecosystems are determined by the river bed's gradient or by the velocity of the current. Faster moving turbulent water typically contains greater concentrations of dissolved oxygen, which supports greater biodiversity than the slow-moving water of pools. These distinctions form the basis for the division of rivers into upland and lowland rivers.

Ecological stoichiometry considers how the balance of energy and elements influences living systems. Similar to chemical stoichiometry, ecological stoichiometry is founded on constraints of mass balance as they apply to organisms and their interactions in ecosystems. Specifically, how does the balance of energy and elements affect and how is this balance affected by organisms and their interactions. Concepts of ecological stoichiometry have a long history in ecology with early references to the constraints of mass balance made by Liebig, Lotka, and Redfield. These earlier concepts have been extended to explicitly link the elemental physiology of organisms to their food web interactions and ecosystem function.
Trophic cascades are powerful indirect interactions that can control entire ecosystems, occurring when a trophic level in a food web is suppressed. For example, a top-down cascade will occur if predators are effective enough in predation to reduce the abundance, or alter the behavior of their prey, thereby releasing the next lower trophic level from predation.

In ecology, a community is a group or association of populations of two or more different species occupying the same geographical area at the same time, also known as a biocoenosis, biotic community, biological community, ecological community, or life assemblage. The term community has a variety of uses. In its simplest form it refers to groups of organisms in a specific place or time, for example, "the fish community of Lake Ontario before industrialization".

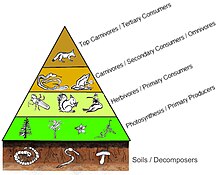
The trophic level of an organism is the position it occupies in a food web. A food chain is a succession of organisms that eat other organisms and may, in turn, be eaten themselves. The trophic level of an organism is the number of steps it is from the start of the chain. A food web starts at trophic level 1 with primary producers such as plants, can move to herbivores at level 2, carnivores at level 3 or higher, and typically finish with apex predators at level 4 or 5. The path along the chain can form either a one-way flow or a food "web". Ecological communities with higher biodiversity form more complex trophic paths.
An ecological network is a representation of the biotic interactions in an ecosystem, in which species (nodes) are connected by pairwise interactions (links). These interactions can be trophic or symbiotic. Ecological networks are used to describe and compare the structures of real ecosystems, while network models are used to investigate the effects of network structure on properties such as ecosystem stability.

An autotroph is an organism that produces complex organic compounds using carbon from simple substances such as carbon dioxide, generally using energy from light (photosynthesis) or inorganic chemical reactions (chemosynthesis). They convert an abiotic source of energy into energy stored in organic compounds, which can be used by other organisms. Autotrophs do not need a living source of carbon or energy and are the producers in a food chain, such as plants on land or algae in water. Autotrophs can reduce carbon dioxide to make organic compounds for biosynthesis and as stored chemical fuel. Most autotrophs use water as the reducing agent, but some can use other hydrogen compounds such as hydrogen sulfide.
In ecology, the term productivity refers to the rate of generation of biomass in an ecosystem, usually expressed in units of mass per volume per unit of time, such as grams per square metre per day. The unit of mass can relate to dry matter or to the mass of generated carbon. The productivity of autotrophs, such as plants, is called primary productivity, while the productivity of heterotrophs, such as animals, is called secondary productivity.

Fishing down the food web is the process whereby fisheries in a given ecosystem, "having depleted the large predatory fish on top of the food web, turn to increasingly smaller species, finally ending up with previously spurned small fish and invertebrates".
A food chain is a linear network of links in a food web, often starting with an autotroph, also called a producer, and typically ending at an apex predator, detritivore, or decomposer. It is not the same as a food web. A food chain depicts relations between species based on what they consume for energy in trophic levels, and they are most commonly quantified in length-the number of links between a trophic consumer and the base of the chain.

Energy, nutrients, and contaminants derived from aquatic ecosystems and transferred to terrestrial ecosystems are termed aquatic-terrestrial subsidies or, more simply, aquatic subsidies. Common examples of aquatic subsidies include organisms that move across habitat boundaries and deposit their nutrients as they decompose in terrestrial habitats or are consumed by terrestrial predators, such as spiders, lizards, birds, and bats. Aquatic insects that develop within streams and lakes before emerging as winged adults and moving to terrestrial habitats contribute to aquatic subsidies. Fish removed from aquatic ecosystems by terrestrial predators are another important example. Conversely, the flow of energy and nutrients from terrestrial ecosystems to aquatic ecosystems are considered terrestrial subsidies; both aquatic subsidies and terrestrial subsidies are types of cross-boundary subsidies. Energy and nutrients are derived from outside the ecosystem where they are ultimately consumed.

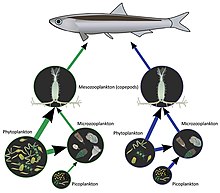
A marine food web is a food web of marine life. At the base of the ocean food web are single-celled algae and other plant-like organisms known as phytoplankton. The second trophic level is occupied by zooplankton which feed off the phytoplankton. Higher order consumers complete the web. There has been increasing recognition in recent years that marine microorganisms.