
Ascomycota is a phylum of the kingdom Fungi that, together with the Basidiomycota, forms the subkingdom Dikarya. Its members are commonly known as the sac fungi or ascomycetes. It is the largest phylum of Fungi, with over 64,000 species. The defining feature of this fungal group is the "ascus", a microscopic sexual structure in which nonmotile spores, called ascospores, are formed. However, some species of the Ascomycota are asexual, meaning that they do not have a sexual cycle and thus do not form asci or ascospores. Familiar examples of sac fungi include morels, truffles, brewers' and bakers' yeast, dead man's fingers, and cup fungi. The fungal symbionts in the majority of lichens such as Cladonia belong to the Ascomycota.

A hypha is a long, branching, filamentous structure of a fungus, oomycete, or actinobacterium. In most fungi, hyphae are the main mode of vegetative growth, and are collectively called a mycelium.

A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, its root system. Mycorrhizae play important roles in plant nutrition, soil biology, and soil chemistry.

Fungus-growing ants comprise all the known fungus-growing ant species participating in ant–fungus mutualism. They are known for cutting grasses and leaves, carrying them to their colonies' nests, and using them to grow fungus on which they later feed.

The ant–fungus mutualism is a symbiosis seen between certain ant and fungal species, in which ants actively cultivate fungus much like humans farm crops as a food source. There is only evidence of two instances in which this form of agriculture evolved in ants resulting in a dependence on fungi for food. These instances were the attine ants and some ants that are part of the Megalomyrmex genus. In some species, the ants and fungi are dependent on each other for survival. This type of codependency is prevalent among herbivores who rely on plant material for nutrition. The fungus’ ability to convert the plant material into a food source accessible to their host makes them the ideal partner. The leafcutter ant is a well-known example of this symbiosis. Leafcutter ants species can be found in southern South America up to the United States. However, ants are not the only ground-dwelling arthropods which have developed symbioses with fungi. A similar mutualism with fungi is also noted in termites within the subfamily Macrotermitinae which are widely distributed throughout the Old World tropics with the highest diversity in Africa.

A sclerotium, is a compact mass of hardened fungal mycelium containing food reserves. One role of sclerotia is to survive environmental extremes. In some higher fungi such as ergot, sclerotia become detached and remain dormant until favorable growth conditions return. Sclerotia initially were mistaken for individual organisms and described as separate species until Louis René Tulasne proved in 1853 that sclerotia are only a stage in the life cycle of some fungi. Further investigation showed that this stage appears in many fungi belonging to many diverse groups. Sclerotia are important in the understanding of the life cycle and reproduction of fungi, as a food source, as medicine, and in agricultural blight management.

The term mycangium is used in biology for special structures on the body of an animal that are adapted for the transport of symbiotic fungi. This is seen in many xylophagous insects, which apparently derive much of their nutrition from the digestion of various fungi that are growing amidst the wood fibers. In some cases, as in ambrosia beetles, the fungi are the sole food, and the excavations in the wood are simply to make a suitable microenvironment for the fungus to grow. In other cases, wood tissue is the main food, and fungi weaken the defense response from the host plant.

Trichoderma viride is a fungus and a biofungicide.

The Macrotermitinae, the fungus-growing termites, constitute a subfamily of the family Termitidae that is only found within the Old World tropics.

A fungus is any member of the group of eukaryotic organisms that includes microorganisms such as yeasts and molds, as well as the more familiar mushrooms. These organisms are classified as a kingdom, separately from the other eukaryotic kingdoms, which, by one traditional classification, includes Plantae, Animalia, Protozoa, and Chromista.

Armillaria gallica is a species of honey mushroom in the family Physalacriaceae of the order Agaricales. The species is a common and ecologically important wood-decay fungus that can live as a saprobe, or as an opportunistic parasite in weakened tree hosts to cause root or butt rot. It is found in temperate regions of Asia, North America, and Europe. The species forms fruit bodies singly or in groups in soil or rotting wood. The fungus has been inadvertently introduced to South Africa. Armillaria gallica has had a confusing taxonomy, due in part to historical difficulties encountered in distinguishing between similar Armillaria species. The fungus received international attention in the early 1990s when an individual colony living in a Michigan forest was reported to cover an area of 15 hectares, weigh at least 9.5 tonnes, and be 1,500 years old. This individual is popularly known as the "humongous fungus", and is a tourist attraction and inspiration for an annual mushroom-themed festival in Crystal Falls. Recent studies have revised the fungus's age to 2,500 years and its size to about 400 tonnes, four times the original estimate.
The mycorrhizosphere is the region around a mycorrhizal fungus in which nutrients released from the fungus increase the microbial population and its activities. The roots of most terrestrial plants, including most crop plants and almost all woody plants, are colonized by mycorrhiza-forming symbiotic fungi. In this relationship, the plant roots are infected by a fungus, but the rest of the fungal mycelium continues to grow through the soil, digesting and absorbing nutrients and water and sharing these with its plant host. The fungus in turn benefits by receiving photosynthetic sugars from its host. The mycorrhizosphere consists of roots, hyphae of the directly connected mycorrhizal fungi, associated microorganisms, and the soil in their direct influence.

An ectomycorrhiza is a form of symbiotic relationship that occurs between a fungal symbiont, or mycobiont, and the roots of various plant species. The mycobiont is often from the phyla Basidiomycota and Ascomycota, and more rarely from the Zygomycota. Ectomycorrhizas form on the roots of around 2% of plant species, usually woody plants, including species from the birch, dipterocarp, myrtle, beech, willow, pine and rose families. Research on ectomycorrhizas is increasingly important in areas such as ecosystem management and restoration, forestry and agriculture.
Fungal-bacterial endosymbiosis encompasses the mutualistic relationship between a fungus and intracellular bacteria species residing within the fungus. Many examples of endosymbiotic relationships between bacteria and plants, algae and insects exist and have been well characterized, however fungal-bacteria endosymbiosis has been less well described.
Orchid mycorrhizae are endomycorrhizal fungi which develop symbiotic relationships with the roots and seeds of plants of the family Orchidaceae. Nearly all orchids are myco-heterotrophic at some point in their life cycle. Orchid mycorrhizae are critically important during orchid germination, as an orchid seed has virtually no energy reserve and obtains its carbon from the fungal symbiont.
A mycoparasite is an organism with the ability to parasitize fungi.

Leucoagaricus gongylophorus is a fungus in the family Agaricaceae which is cultivated by certain leafcutter ants. Like other species of fungi cultivated by ants, L. gongylophorus produces gongylidia, nutrient-rich hyphal swellings upon which the ants feed. Production of mushrooms occurs only once ants abandon the nest. L. gongylophorus is farmed by leaf cutter ant species belonging to the genera Atta and Acromyrmex, amongst others.
Hemibiotrophs are the spectrum of plant pathogens, including bacteria, oomycete and a group of plant pathogenic fungi that keep its host alive while establishing itself within the host tissue, taking up the nutrients with brief biotrophic-like phase. It then, in later stages of infection switches to a necrotrophic life-style, where it rampantly kills the host cells, deriving its nutrients from the dead tissues.
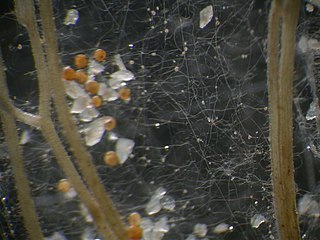
Gigaspora margarita is an Arbuscular Mycorrhizal Fungi (AMF) which means it is an obligate symbiont that creates mutualistic relationships with many different plant species. Being an AMF, G. margarita does not produce a fruiting body. All of its mycelium will be found in the soil, associating with plant roots. Though hard to distinguish between different species of AMF, microscopic distinctions can be made. A prominent morphological distinction for species in the Gigasporaceae family is their large sized spores. Gigaspora margarita is characterized by its large, white, pearl-like spores found anywhere from 260 - 400 micrometers. This is where it gets its name as margarita in Latin means pearl.

Trichoderma atroviride is a filamentous fungal species commonly found in the soil. This fungal species is of particular interest to researchers due to the plethora of secondary metabolites it makes which are used in industry The genus Trichoderma is known for its ubiquity in almost all soils and being easy to culture. Many Trichoderma's are also avirulent plant symbionts.






