
A conidium, sometimes termed an asexual chlamydospore or chlamydoconidium, is an asexual, non-motile spore of a fungus. The word conidium comes from the Ancient Greek word for dust, κόνις (kónis). They are also called mitospores due to the way they are generated through the cellular process of mitosis. They are produced exogenously. The two new haploid cells are genetically identical to the haploid parent, and can develop into new organisms if conditions are favorable, and serve in biological dispersal.

An appressorium is a specialized cell typical of many fungal plant pathogens that is used to infect host plants. It is a flattened, hyphal "pressing" organ, from which a minute infection peg grows and enters the host, using turgor pressure capable of punching through even Mylar.

Rhizoctonia solani is a species of fungus in the order Cantharellales. Basidiocarps are thin, effused, and web-like, but the fungus is more typically encountered in its anamorphic state, as hyphae and sclerotia. The name Rhizoctonia solani is currently applied to a complex of related species that await further research. In its wide sense, Rhizoctonia solani is a facultative plant pathogen with a wide host range and worldwide distribution. It causes various plant diseases such as root rot, damping off, and wire stem. It can also form mycorrhizal associations with orchids.
Glomerella graminicola is an economically important crop parasite affecting both wheat and maize where it causes the plant disease Anthracnose Leaf Blight.

Thielaviopsis basicola is the plant-pathogen fungus responsible for black root rot disease. This particular disease has a large host range, affecting woody ornamentals, herbaceous ornamentals, agronomic crops, and even vegetable crops. Examples of susceptible hosts include petunia, pansy, poinsettia, tobacco, cotton, carrot, lettuce, tomato, and others. Symptoms of this disease resemble nutrient deficiency but are truly a result of the decaying root systems of plants. Common symptoms include chlorotic lower foliage, yellowing of plant, stunting or wilting, and black lesions along the roots. The lesions along the roots may appear red at first, getting darker and turning black as the disease progresses. Black root lesions that begin in the middle of a root can also spread further along the roots in either direction. Due to the nature of the pathogen, the disease can easily be identified by the black lesions along the roots, especially when compared to healthy roots. The black lesions that appear along the roots are a result of the formation of chlamydospores, resting spores of the fungus that contribute to its pathogenicity. The chlamydospores are a dark brown-black color and cause the "discoloration" of the roots when they are produced in large amounts.

Colletotrichum kahawae is a fungal plant pathogen that causes coffee berry disease (CBD) on Coffea arabica crops. The pathogen is an ascomycete that reproduces asexually. The asexual spores (conidia) are stored within acervuli. This disease is considered to be one of the major factors hampering C.arabica production in the African continent, which represents the current geographic range of the fungus. Coffee berry disease causes dark necrosis in spots and causes the green berries of the coffee to drop prematurely. High humidity, relatively warm temperatures, and high altitude are ideal for disease formation. Given the severity of the disease and the lack of effective control measures, there is great concern that the fungus may spread to other coffee producing continents, such as South America, which could have catastrophic consequences.
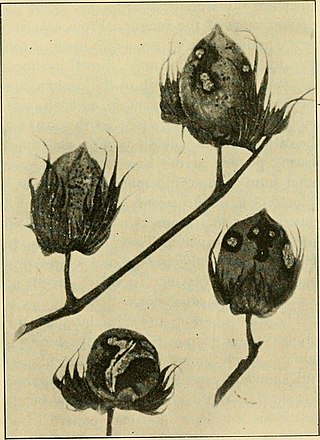
Colletotrichum gossypii is a plant pathogen. This fungus is affiliated with cotton plants where it causes anthracnose. Its reproduction in the plants is asexual. The conidia have only one nucleus. Before conidia germination fusion by mean of conidial anastomosis tube could happen. The conidia could germinate in media plates.
Alternaria carthami is a necrotrophic plant pathogen of safflower. The fungus is in the order Pleosporales and family Pleosporaceae. It was first isolated in India, has spread globally and can have devastating effects on safflower yield, and resultant oilseed production. A. carthami is known to be seed-borne and appears as irregular brown lesions on safflower leaves and stems.

Phomopsis obscurans is a common fungus found in strawberry plants, which causes the disease of leaf blight. Common symptoms caused by the pathogen begin as small circular reddish-purple spots and enlarge to form V-shaped lesions that follow the vasculature of the plant's leaves. Although the fungus infects leaves early in the growing season when the plants are beginning to develop, leaf blight symptoms are most apparent on older plants towards the end of the growing season. The disease can weaken strawberry plants through the destruction of foliage, which results in reduced yields. In years highly favorable for disease development, leaf blight can ultimately lead to the death of the strawberry plants. A favorable environment for the growth and development of the Phomopsis obscurans pathogen is that of high temperature, high inoculum density, a long period of exposure to moisture, and immature host tissue. In the case of disease management, a conjunction of cultural practices is the most effective way of reducing the infection.
Colletotrichum capsici is a species of fungus and plant pathogen which causes leaf blight on Chlorophytum borivilianum, basil, chickpea and pepper as well as dieback in pigeonpea and anthracnose in poinsettia.

Colletotrichum coccodes is a plant pathogen, which causes anthracnose on tomato and black dot disease of potato. Fungi survive on crop debris and disease emergence is favored by warm temperatures and wet weather.

Glomerella cingulata is a fungal plant pathogen, being the name of the sexual stage (teleomorph) while the more commonly referred to asexual stage (anamorph) is called Colletotrichum gloeosporioides. For most of this article the pathogen will be referred to as C. gloeosporioides. This pathogen is a significant problem worldwide, causing anthracnose and fruit rotting diseases on hundreds of economically important hosts.
Microdochium panattonianum is a fungal plant pathogen. This pathogen causes anthracnose of lettuce, a disease which produces necrotic lesions in cultivated lettuce. In extended periods of wet weather, M. panattonianum can cause significant crop-losses. The impact of this pathogen is exacerbated by farming lettuce without crop rotation, and by planting of susceptible lettuce varieties, such as Romaine lettuce.
Colletotrichum fragariae is a fungal plant pathogen infecting strawberries. It is not a well known fungus, and there are many similar fungi that are related to it. It is part of the Colletotrichum genus. It is a pathogen that occurs in strawberries. It leads to the disease known as anthracnose. This is typically at the crown of the strawberry, which is why it is often called crown rot. It is also known as the Anthracnose Crown rot. The fungus also infects leaves and is known as leaf spot, which is common among all Colletotrichum. This is not as common in the fragariae, as it is more common in the crown. This fungus is also better at infecting younger strawberries/seedlings. The most common way to control this disease is fungicides that are harmful to the environment. There have been studies done to see if the fungus infects other hosts but other than some weeds, it is very specific to Strawberries.

Botrytis fabae is a plant pathogen, a fungus that causes chocolate spot disease of broad or fava bean plants, Vicia faba. It was described scientifically by Mexican-born Galician microbiologist Juan Rodríguez Sardiña in 1929.
Conidial anastomosis tubes (CATs) are cells formed from the conidia of many filamentous fungi. These cells have a tubular shape and form an anastomosis (bridge) that allows fusion between conidia.
Entomophaga grylli is a fungal pathogen which infects and kills grasshoppers. It is the causal agent of one of the most widespread diseases affecting grasshoppers. This is sometimes known as summit disease because infected insects climb to the upper part of a plant and grip the tip of the stem as they die; this ensures widespread dispersal of the fungal spores. The fungus is a species complex with several different pathotypes, each one of which seems to be host-specific to different subfamilies of grasshoppers. The pathogen is being investigated for its possible use in biological pest control of grasshoppers.

Alternaria brassicicola is a fungal necrotrophic plant pathogen that causes black spot disease on a wide range of hosts, particularly in the genus of Brassica, including a number of economically important crops such as cabbage, Chinese cabbage, cauliflower, oilseeds, broccoli and canola. Although mainly known as a significant plant pathogen, it also contributes to various respiratory allergic conditions such as asthma and rhinoconjunctivitis. Despite the presence of mating genes, no sexual reproductive stage has been reported for this fungus. In terms of geography, it is most likely to be found in tropical and sub-tropical regions, but also in places with high rain and humidity such as Poland. It has also been found in Taiwan and Israel. Its main mode of propagation is vegetative. The resulting conidia reside in the soil, air and water. These spores are extremely resilient and can overwinter on crop debris and overwintering herbaceous plants.
Hemibiotrophs are the spectrum of plant pathogens, including bacteria, oomycete and a group of plant pathogenic fungi that keep its host alive while establishing itself within the host tissue, taking up the nutrients with brief biotrophic-like phase. It then, in later stages of infection switches to a necrotrophic life-style, where it rampantly kills the host cells, deriving its nutrients from the dead tissues.

Colletotrichum fioriniae is a fungal plant pathogen and endophyte of fruits and foliage of many broadleaved plants worldwide. It causes diseases on agriculturally important crops, including anthracnose of strawberry, ripe rot of grapes, bitter rot of apple, anthracnose of peach, and anthracnose of blueberry. Its ecological role in the natural environment is less well understood, other than it is a common leaf endophyte of many temperate trees and shrubs and in some cases may function as an entomopathogen.



