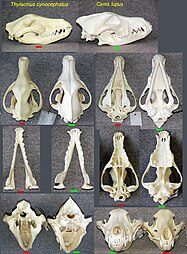
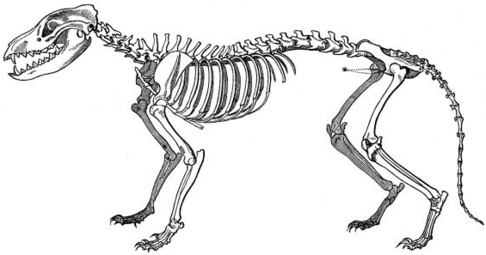
Common descent is a concept in evolutionary biology applicable when one species is the ancestor of two or more species later in time. According to modern evolutionary biology, all living beings could be descendants of a unique ancestor commonly referred to as the last universal common ancestor (LUCA) of all life on Earth.
Genomic imprinting is an epigenetic phenomenon that causes genes to be expressed or not, depending on whether they are inherited from the female or male parent. Genes can also be partially imprinted. Partial imprinting occurs when alleles from both parents are differently expressed rather than complete expression and complete suppression of one parent's allele. Forms of genomic imprinting have been demonstrated in fungi, plants and animals. In 2014, there were about 150 imprinted genes known in mice and about half that in humans. As of 2019, 260 imprinted genes have been reported in mice and 228 in humans.
Macroevolution usually means the evolution of large-scale structures and traits that go significantly beyond the intraspecific variation found in microevolution. In other words, macroevolution is the evolution of taxa above the species level.
Molecular evolution is the process of change in the sequence composition of cellular molecules such as DNA, RNA, and proteins across generations. The field of molecular evolution uses principles of evolutionary biology and population genetics to explain patterns in these changes. Major topics in molecular evolution concern the rates and impacts of single nucleotide changes, neutral evolution vs. natural selection, origins of new genes, the genetic nature of complex traits, the genetic basis of speciation, the evolution of development, and ways that evolutionary forces influence genomic and phenotypic changes.
The molecular clock is a figurative term for a technique that uses the mutation rate of biomolecules to deduce the time in prehistory when two or more life forms diverged. The biomolecular data used for such calculations are usually nucleotide sequences for DNA, RNA, or amino acid sequences for proteins.

Afrotheria is a superorder of mammals, the living members of which belong to groups that are either currently living in Africa or of African origin: golden moles, elephant shrews, otter shrews, tenrecs, aardvarks, hyraxes, elephants, sea cows, and several extinct clades. Most groups of afrotheres share little or no superficial resemblance, and their similarities have only become known in recent times because of genetics and molecular studies. Many afrothere groups are found mostly or exclusively in Africa, reflecting the fact that Africa was an island continent from the Cretaceous until the early Miocene around 20 million years ago, when Afro-Arabia collided with Eurasia.
Thermogenesis is the process of heat production in organisms. It occurs in all warm-blooded animals, and also in a few species of thermogenic plants such as the Eastern skunk cabbage, the Voodoo lily, and the giant water lilies of the genus Victoria. The lodgepole pine dwarf mistletoe, Arceuthobium americanum, disperses its seeds explosively through thermogenesis.
Evolvability is defined as the capacity of a system for adaptive evolution. Evolvability is the ability of a population of organisms to not merely generate genetic diversity, but to generate adaptive genetic diversity, and thereby evolve through natural selection.
Parallel evolution is the similar development of a trait in distinct species that are not closely related, but share a similar original trait in response to similar evolutionary pressure.
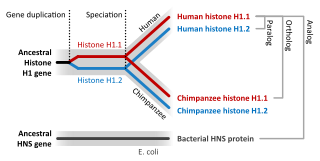
Sequence homology is the biological homology between DNA, RNA, or protein sequences, defined in terms of shared ancestry in the evolutionary history of life. Two segments of DNA can have shared ancestry because of three phenomena: either a speciation event (orthologs), or a duplication event (paralogs), or else a horizontal gene transfer event (xenologs).

Tyrosinase is an oxidase that is the rate-limiting enzyme for controlling the production of melanin. The enzyme is mainly involved in two distinct reactions of melanin synthesis otherwise known as the Raper Mason pathway. Firstly, the hydroxylation of a monophenol and secondly, the conversion of an o-diphenol to the corresponding o-quinone. o-Quinone undergoes several reactions to eventually form melanin. Tyrosinase is a copper-containing enzyme present in plant and animal tissues that catalyzes the production of melanin and other pigments from tyrosine by oxidation. It is found inside melanosomes which are synthesized in the skin melanocytes. In humans, the tyrosinase enzyme is encoded by the TYR gene.

Divergent evolution or divergent selection is the accumulation of differences between closely related populations within a species, sometimes leading to speciation. Divergent evolution is typically exhibited when two populations become separated by a geographic barrier and experience different selective pressures that cause adaptations. After many generations and continual evolution, the populations become less able to interbreed with one another. The American naturalist J. T. Gulick (1832–1923) was the first to use the term "divergent evolution", with its use becoming widespread in modern evolutionary literature. Examples of divergence in nature are the adaptive radiation of the finches of the Galápagos, changes in mobbing behavior of the kittiwake, and the evolution of the modern-day dog from the wolf.
Evidence of common descent of living organisms has been discovered by scientists researching in a variety of disciplines over many decades, demonstrating that all life on Earth comes from a single ancestor. This forms an important part of the evidence on which evolutionary theory rests, demonstrates that evolution does occur, and illustrates the processes that created Earth's biodiversity. It supports the modern evolutionary synthesis—the current scientific theory that explains how and why life changes over time. Evolutionary biologists document evidence of common descent, all the way back to the last universal common ancestor, by developing testable predictions, testing hypotheses, and constructing theories that illustrate and describe its causes.

A catalytic triad is a set of three coordinated amino acids that can be found in the active site of some enzymes. Catalytic triads are most commonly found in hydrolase and transferase enzymes. An acid-base-nucleophile triad is a common motif for generating a nucleophilic residue for covalent catalysis. The residues form a charge-relay network to polarise and activate the nucleophile, which attacks the substrate, forming a covalent intermediate which is then hydrolysed to release the product and regenerate free enzyme. The nucleophile is most commonly a serine or cysteine amino acid, but occasionally threonine or even selenocysteine. The 3D structure of the enzyme brings together the triad residues in a precise orientation, even though they may be far apart in the sequence.

Why Evolution is True is a popular science book by American biologist Jerry Coyne. It was published in 2009, dubbed "Darwin Year" as it marked the bicentennial of Charles Darwin and the hundred and fiftieth anniversary of the publication of his On the Origin of Species By Means of Natural Selection. Coyne examines the evidence for evolution, some of which was known to Darwin (biogeography) and some of which has emerged in recent years. The book was a New York Times bestseller, and reviewers praised the logic of Coyne's arguments and the clarity of his prose. It was reprinted as part of the Oxford Landmark Science series.
Enzyme promiscuity is the ability of an enzyme to catalyse a fortuitous side reaction in addition to its main reaction. Although enzymes are remarkably specific catalysts, they can often perform side reactions in addition to their main, native catalytic activity. These promiscuous activities are usually slow relative to the main activity and are under neutral selection. Despite ordinarily being physiologically irrelevant, under new selective pressures these activities may confer a fitness benefit therefore prompting the evolution of the formerly promiscuous activity to become the new main activity. An example of this is the atrazine chlorohydrolase from Pseudomonas sp. ADP that evolved from melamine deaminase, which has very small promiscuous activity toward atrazine, a man-made chemical.
A protein superfamily is the largest grouping (clade) of proteins for which common ancestry can be inferred. Usually this common ancestry is inferred from structural alignment and mechanistic similarity, even if no sequence similarity is evident. Sequence homology can then be deduced even if not apparent. Superfamilies typically contain several protein families which show sequence similarity within each family. The term protein clan is commonly used for protease and glycosyl hydrolases superfamilies based on the MEROPS and CAZy classification systems.
Recurrent evolution is the repeated evolution of a particular trait, character, or mutation. Most evolution is the result of drift, often interpreted as the random chance of some alleles being passed down to the next generation and others not. Recurrent evolution is said to occur when patterns emerge from this stochastic process when looking across multiple distinct populations. These patterns are of particular interest to evolutionary biologists, as they can demonstrate the underlying forces governing evolution.
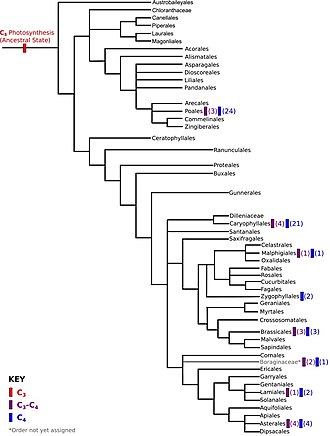
The evolution of photosynthesis refers to the origin and subsequent evolution of photosynthesis, the process by which light energy is used to assemble sugars from carbon dioxide and a hydrogen and electron source such as water. The process of photosynthesis was discovered by Jan Ingenhousz, a Dutch-born British physician and scientist, first publishing about it in 1779.