Related Research Articles

Ecology is the natural science of the relationships among living organisms, including humans, and their physical environment. Ecology considers organisms at the individual, population, community, ecosystem, and biosphere level. Ecology overlaps with the closely related sciences of biogeography, evolutionary biology, genetics, ethology, and natural history.

Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.
A complex system is a system composed of many components which may interact with each other. Examples of complex systems are Earth's global climate, organisms, the human brain, infrastructure such as power grid, transportation or communication systems, complex software and electronic systems, social and economic organizations, an ecosystem, a living cell, and ultimately the entire universe.

Mutualism describes the ecological interaction between two or more species where each species has a net benefit. Mutualism is a common type of ecological interaction, one that can come from a parasitic interaction. Prominent examples include most vascular plants engaged in mutualistic interactions with mycorrhizae, flowering plants being pollinated by animals, vascular plants being dispersed by animals, and corals with zooxanthellae, among many others. Mutualism can be contrasted with interspecific competition, in which each species experiences reduced fitness, and exploitation, or parasitism, in which one species benefits at the expense of the other.
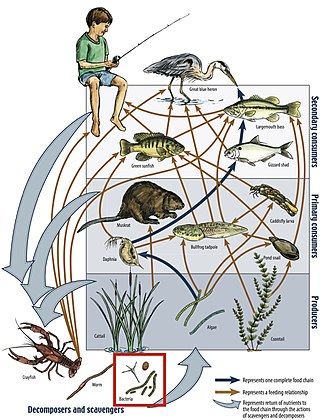
A food web is the natural interconnection of food chains and a graphical representation of what-eats-what in an ecological community. Ecologists can broadly define all life forms as either autotrophs or heterotrophs, based on their trophic levels, the position that they occupy in the food web. To maintain their bodies, grow, develop, and to reproduce, autotrophs produce organic matter from inorganic substances, including both minerals and gases such as carbon dioxide. These chemical reactions require energy, which mainly comes from the Sun and largely by photosynthesis, although a very small amount comes from bioelectrogenesis in wetlands, and mineral electron donors in hydrothermal vents and hot springs. These trophic levels are not binary, but form a gradient that includes complete autotrophs, which obtain their sole source of carbon from the atmosphere, mixotrophs, which are autotrophic organisms that partially obtain organic matter from sources other than the atmosphere, and complete heterotrophs that must feed to obtain organic matter.

In ecology, a biological interaction is the effect that a pair of organisms living together in a community have on each other. They can be either of the same species, or of different species. These effects may be short-term, or long-term, both often strongly influence the adaptation and evolution of the species involved. Biological interactions range from mutualism, beneficial to both partners, to competition, harmful to both partners. Interactions can be direct when physical contact is established or indirect, through intermediaries such as shared resources, territories, ecological services, metabolic waste, toxins or growth inhibitors. This type of relationship can be shown by net effect based on individual effects on both organisms arising out of relationship.

The soil food web is the community of organisms living all or part of their lives in the soil. It describes a complex living system in the soil and how it interacts with the environment, plants, and animals.
In ecology, an ecosystem is said to possess ecological stability if it is capable of returning to its equilibrium state after a perturbation or does not experience unexpected large changes in its characteristics across time. Although the terms community stability and ecological stability are sometimes used interchangeably, community stability refers only to the characteristics of communities. It is possible for an ecosystem or a community to be stable in some of their properties and unstable in others. For example, a vegetation community in response to a drought might conserve biomass but lose biodiversity.
Nestedness is a measure of structure in an ecological system, usually applied to species-sites systems, or species-species interaction networks.
Soil ecology is the study of the interactions among soil organisms, and between biotic and abiotic aspects of the soil environment. It is particularly concerned with the cycling of nutrients, formation and stabilization of the pore structure, the spread and vitality of pathogens, and the biodiversity of this rich biological community.

In ecology, a community is a group or association of populations of two or more different species occupying the same geographical area at the same time, also known as a biocoenosis, biotic community, biological community, ecological community, or life assemblage. The term community has a variety of uses. In its simplest form it refers to groups of organisms in a specific place or time, for example, "the fish community of Lake Ontario before industrialization".
A biological network is a method of representing systems as complex sets of binary interactions or relations between various biological entities. In general, networks or graphs are used to capture relationships between entities or objects. A typical graphing representation consists of a set of nodes connected by edges.
A food chain is a linear network of links in a food web starting from producer organisms and ending at an apex predator species, detritivores, or decomposer species. A food chain also shows how organisms are related to each other by the food they eat. Each level of a food chain represents a different trophic level. A food chain differs from a food web because the complex network of different animals' feeding relations are aggregated and the chain only follows a direct, linear pathway of one animal at a time. Natural interconnections between food chains make it a food web.

Evolving digital ecological networks are webs of interacting, self-replicating, and evolving computer programs that experience the same major ecological interactions as biological organisms. Despite being computational, these programs evolve quickly in an open-ended way, and starting from only one or two ancestral organisms, the formation of ecological networks can be observed in real-time by tracking interactions between the constantly evolving organism phenotypes. These phenotypes may be defined by combinations of logical computations that digital organisms perform and by expressed behaviors that have evolved. The types and outcomes of interactions between phenotypes are determined by task overlap for logic-defined phenotypes and by responses to encounters in the case of behavioral phenotypes. Biologists use these evolving networks to study active and fundamental topics within evolutionary ecology.

A pollination network is a bipartite mutualistic network in which plants and pollinators are the nodes, and the pollination interactions form the links between these nodes. The pollination network is bipartite as interactions only exist between two distinct, non-overlapping sets of species, but not within the set: a pollinator can never be pollinated, unlike in a predator-prey network where a predator can be depredated. A pollination network is two-modal, i.e., it includes only links connecting plant and animal communities.

A Mesopredator is a predator that occupies a mid-ranking trophic level in a food web. There is no standard definition of a mesopredator, but mesopredators are usually medium-sized carnivorous or omnivorous animals, such as raccoons, foxes, or coyotes. They are often defined by contrast from apex predators or prey in a particular food web. Mesopredators typically prey on smaller animals.
Eric L. Berlow is an American ecologist and data scientist. He co-founded a visual data interface company, which was acquired by Rakuten Inc. in 2016. He now runs Vibrant Data Labs, a social impact data science group currently focused the building on an open-source framework for tracking the flows of money to climate mitigation and resilience efforts on the ground. Prior to Vibrant Data Labs, Berlow was the founding director of the University of California's first science and education institute inside Yosemite National Park which facilitated efforts to leverage data for informing conservation policy and natural resource management. Berlow is internationally recognized for his research on ecological complexity, with articles in Nature, Science, and Proceedings of the National Academy of Sciences. He is best known for his TED talks on simplifying complexity and finding hidden patterns in complex data. Berlow has received a TED Fellowship, a TED Senior Fellowship, an Alexander Von Humboldt Fellowship, a National Science Foundation Post-doctoral Fellowship, and a National Center for Ecological Analysis and Synthesis Fellowship. He was named one of the top 100 Creatives by Origin magazine.

Pedro Diego Jordano Barbudo is an ecologist, conservationist, researcher, focused on evolutionary ecology and ecological interactions. He is an honorary professor and associate professor at University of Sevilla, Spain. Most of his fieldwork is done in Parque Natural de las Sierras de Cazorla, Segura y Las Villas, in the eastern side of Andalucia, and in Doñana National Park, where he holds the title of Research Professor for the Estación Biológica Doñana, Spanish Council for Scientific Research (CSIC). Since 2000 he has been actively doing research in Brazil, with fieldwork in the SE Atlantic rainforest.
Jennifer Dunne is an American ecologist whose research focuses on the network structure of food webs. One of 14 scientists who led critical advances in food web research over the last century, according to the journal Food Webs, Dunne uses ecological network research to compare the varying ways humans interact with other species through space and time, providing a quantitative perspective on sustainability of socio-ecological systems.
Jordi Bascompte is a professor of ecology at the University of Zurich and the director of its specialized master's program on quantitative environmental sciences. He is best known for having brought the interactions of mutual benefit between plants and animals into community ecology, at the time largely dominated by predation and competition. His application of network theory to the study of mutualism has identified general laws that determine the way in which species interactions shape biodiversity.
References
Specific
- Bascompte, J.; Jordano, P.; Melian, C.J.; Olesen, J.M. (2003). "The nested assembly of plant-animal mutualistic networks". Proceedings of the National Academy of Sciences. 100 (16): 9383–9387. Bibcode:2003PNAS..100.9383B. doi: 10.1073/pnas.1633576100 . PMC 170927 . PMID 12881488.
- Burgos, E.; Ceva, H.; Perazzo, R.P.J.; Devoto, M.; Medan, D.; Zimmermann, M.; Delbue, A.M. (2007). "Why nestedness in mutualistic networks?". Journal of Theoretical Biology. 249 (2): 307–313. arXiv: q-bio/0609005 . Bibcode:2007JThBi.249..307B. doi:10.1016/j.jtbi.2007.07.030. PMID 17897679. S2CID 10258963.
- Dunne, J.A.; Williams, R.J.; Martinez, N.D. (2002). "Network structure and biodiversity loss in food webs: robustness increases with connectance". Ecology Letters. 5 (4): 558–567. Bibcode:2002EcolL...5..558D. doi:10.1046/j.1461-0248.2002.00354.x. S2CID 2114852.
- Dunne, J.A.; Williams, R.J.; Martinez, N.D. (2002). "Food-web structure and network theory: The role of connectance and size". Proceedings of the National Academy of Sciences. 99 (20): 12917–12922. Bibcode:2002PNAS...9912917D. doi: 10.1073/pnas.192407699 . PMC 130560 . PMID 12235364.
- Krause, A.E.; Frank, K.A.; Mason, D.M.; Ulanowicz, R.E.; Taylor, W.W. (2003). "Compartments revealed in food-web structure" (PDF). Nature. 426 (6964): 282–285. Bibcode:2003Natur.426..282K. doi:10.1038/nature02115. hdl: 2027.42/62960 . PMID 14628050. S2CID 1752696.
- Memmot, J.; Waser, N.M.; Price, M.V. (2004). "Tolerance of pollination networks to species extinctions". Proceedings of the Royal Society of London B. 271 (1557): 2605–2611. doi:10.1098/rspb.2004.2909. PMC 1691904 . PMID 15615687.
- Okuyama, T.; Holland, J.N. (2008). "Network structure properties mediate the stability of mutualistic communities" (PDF). Ecology Letters. 11 (3): 208–216. Bibcode:2008EcolL..11..208O. doi:10.1111/j.1461-0248.2007.01137.x. PMID 18070101.
- Pimm, S.L. (1984). "The complexity and stability of ecosystems". Nature. 307 (5949): 321–326. Bibcode:1984Natur.307..321P. doi:10.1038/307321a0. S2CID 4317192.
- Reuman, D.C.; Cohen, J.E. (2004). "Trophic links' length and slope in the Tuesday Lake food web with species' body mass and numerical abundance". Journal of Animal Ecology. 73 (5): 852–866. Bibcode:2004JAnEc..73..852R. doi: 10.1111/j.0021-8790.2004.00856.x .
- Schmid-Araya, J.M.; Schmid, P.E.; Robertson, A.; Winterbottom, J.; Gjerlov, C.; Hildrew, A.G. (2002). "Connectance in stream food webs". Journal of Animal Ecology. 71 (6): 1056–1062. Bibcode:2002JAnEc..71.1056S. doi: 10.1046/j.1365-2656.2002.00668.x .
- Stouffer, D.B. (2010). "Scaling from individuals to networks in food webs". Functional Ecology. 24 (1): 44–51. Bibcode:2010FuEco..24...44S. doi: 10.1111/j.1365-2435.2009.01644.x .
- Sole, R.V.; Montoya, J.M. (2001). "Complexity and fragility in ecological networks". Proceedings of the Royal Society of London B. 268 (1480): 2039–2045. arXiv: cond-mat/0011196 . doi:10.1098/rspb.2001.1767. PMC 1088846 . PMID 11571051.
- Vazquez, D.P.; Melian, C.J.; Williams, N.M.; Bluthgen, N.; Krasnov, B.R.; Poulin, R. (2007). "Species abundance and asymmetric interaction strength in ecological networks". Oikos. 116 (7): 1120–1127. CiteSeerX 10.1.1.567.1820 . doi:10.1111/j.2007.0030-1299.15828.x (inactive 2024-02-07).
{{cite journal}}: CS1 maint: DOI inactive as of February 2024 (link) - Williams, R.J.; Berlow, E.L.; Dunne, J.A.; Barabasi, A.L.; Martinez, N.D. (2002). "Two degrees of separation in complex food webs". Proceedings of the National Academy of Sciences. 99 (20): 12913–12916. Bibcode:2002PNAS...9912913W. doi: 10.1073/pnas.192448799 . PMC 130559 . PMID 12235367.
- Zhang, F.; Hui, C.; Terblanche, J.S. (2011). "An interaction switch predicts the nested architecture of mutualistic networks". Ecology Letters. 14 (8): 797–803. Bibcode:2011EcolL..14..797Z. doi:10.1111/j.1461-0248.2011.01647.x. hdl: 10019.1/118286 . PMID 21707903.
- Suweis, S.; Simini, F.; Banavar, J; Maritan, A. (2013). "Emergence of structural and dynamical properties of ecological mutualistic networks". Nature. 500 (7463): 449–452. arXiv: 1308.4807 . Bibcode:2013Natur.500..449S. doi:10.1038/nature12438. PMID 23969462. S2CID 4412384.
- Lever, J. J.; Nes, E. H.; Scheffer, M.; Bascompte, J. (2014). "The sudden collapse of pollinator communities". Ecology Letters. 17 (3): 350–359. Bibcode:2014EcolL..17..350L. doi:10.1111/ele.12236. hdl: 10261/91808 . PMID 24386999.
General
- Bascompte, J (2007). "Networks in ecology". Basic and Applied Ecology. 8 (6): 485–490. Bibcode:2007BApEc...8..485B. doi:10.1016/j.baae.2007.06.003. hdl: 10261/40172 .
- Montoya, J.M.; Pimm, S.L.; Sole, R.V. (2006). "Ecological networks and their fragility". Nature. 442 (7100): 259–264. Bibcode:2006Natur.442..259M. doi:10.1038/nature04927. PMID 16855581. S2CID 592403.