The human genome is a complete set of nucleic acid sequences for humans, encoded as DNA within the 23 chromosome pairs in cell nuclei and in a small DNA molecule found within individual mitochondria. These are usually treated separately as the nuclear genome and the mitochondrial genome. Human genomes include both protein-coding DNA sequences and various types of DNA that does not encode proteins. The latter is a diverse category that includes DNA coding for non-translated RNA, such as that for ribosomal RNA, transfer RNA, ribozymes, small nuclear RNAs, and several types of regulatory RNAs. It also includes promoters and their associated gene-regulatory elements, DNA playing structural and replicatory roles, such as scaffolding regions, telomeres, centromeres, and origins of replication, plus large numbers of transposable elements, inserted viral DNA, non-functional pseudogenes and simple, highly repetitive sequences. Introns make up a large percentage of non-coding DNA. Some of this non-coding DNA is non-functional junk DNA, such as pseudogenes, but there is no firm consensus on the total amount of junk DNA.
Molecular evolution is the process of change in the sequence composition of cellular molecules such as DNA, RNA, and proteins across generations. The field of molecular evolution uses principles of evolutionary biology and population genetics to explain patterns in these changes. Major topics in molecular evolution concern the rates and impacts of single nucleotide changes, neutral evolution vs. natural selection, origins of new genes, the genetic nature of complex traits, the genetic basis of speciation, the evolution of development, and ways that evolutionary forces influence genomic and phenotypic changes.

Forkhead box protein P2 (FOXP2) is a protein that, in humans, is encoded by the FOXP2 gene. FOXP2 is a member of the forkhead box family of transcription factors, proteins that regulate gene expression by binding to DNA. It is expressed in the brain, heart, lungs and digestive system.

Pseudogenes are nonfunctional segments of DNA that resemble functional genes. Most arise as superfluous copies of functional genes, either directly by gene duplication or indirectly by reverse transcription of an mRNA transcript. Pseudogenes are usually identified when genome sequence analysis finds gene-like sequences that lack regulatory sequences needed for transcription or translation, or whose coding sequences are obviously defective due to frameshifts or premature stop codons. Pseudogenes are a type of junk DNA.

A gene family is a set of several similar genes, formed by duplication of a single original gene, and generally with similar biochemical functions. One such family are the genes for human hemoglobin subunits; the ten genes are in two clusters on different chromosomes, called the α-globin and β-globin loci. These two gene clusters are thought to have arisen as a result of a precursor gene being duplicated approximately 500 million years ago.
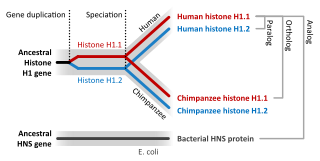
Sequence homology is the biological homology between DNA, RNA, or protein sequences, defined in terms of shared ancestry in the evolutionary history of life. Two segments of DNA can have shared ancestry because of three phenomena: either a speciation event (orthologs), or a duplication event (paralogs), or else a horizontal gene transfer event (xenologs).

The Hominini form a taxonomic tribe of the subfamily Homininae ("hominines"). Hominini includes the extant genera Homo (humans) and Pan and in standard usage excludes the genus Gorilla (gorillas).

Copy number variation (CNV) is a phenomenon in which sections of the genome are repeated and the number of repeats in the genome varies between individuals. Copy number variation is a type of structural variation: specifically, it is a type of duplication or deletion event that affects a considerable number of base pairs. Approximately two-thirds of the entire human genome may be composed of repeats and 4.8–9.5% of the human genome can be classified as copy number variations. In mammals, copy number variations play an important role in generating necessary variation in the population as well as disease phenotype.
Human evolutionary genetics studies how one human genome differs from another human genome, the evolutionary past that gave rise to the human genome, and its current effects. Differences between genomes have anthropological, medical, historical and forensic implications and applications. Genetic data can provide important insights into human evolution.
The Olduvai domain, known until 2018 as DUF1220 and the NBPF repeat, is a protein domain that shows a striking human lineage-specific (HLS) increase in copy number and appears to be involved in human brain evolution. The protein domain has also been linked to several neurogenetic disorders such as schizophrenia and increased severity of autism. In 2018, it was named by its discoverers after Olduvai Gorge in Tanzania, one of the most important archaeological sites for early humans, to reflect data indicating its role in human brain size and evolution.

Cadherin EGF LAG seven-pass G-type receptor 1 also known as flamingo homolog 2 or cadherin family member 9 is a protein that in humans is encoded by the CELSR1 gene.

Protocadherin 11 X-linked, also known as PCDH11X, is a protein which in humans is encoded by the PCDH11X gene.
Forkhead box protein K2 is a protein that in humans is encoded by the FOXK2 gene.

Protocadherin-17 is a protein that in humans is encoded by the PCDH17 gene.

Forkhead box protein D4 is a protein that in humans is encoded by the FOXD4 gene.

Eukaryotic translation initiation factor 1A, Y-chromosomal is a protein that in humans is encoded by the EIF1AY gene.

Methylmalonic aciduria and homocystinuria type D protein, mitochondrial also known as MMADHC is a protein that in humans is encoded by the MMADHC gene.
The chimpanzee–human last common ancestor (CHLCA) is the last common ancestor shared by the extant Homo (human) and Pan genera of Hominini. Estimates of the divergence date vary widely from thirteen to five million years ago.
Cognitive genomics is the sub-field of genomics pertaining to cognitive function in which the genes and non-coding sequences of an organism's genome related to the health and activity of the brain are studied. By applying comparative genomics, the genomes of multiple species are compared in order to identify genetic and phenotypical differences between species. Observed phenotypical characteristics related to the neurological function include behavior, personality, neuroanatomy, and neuropathology. The theory behind cognitive genomics is based on elements of genetics, evolutionary biology, molecular biology, cognitive psychology, behavioral psychology, and neurophysiology.
The myth of the one percent refers to the 1975 study done by Wilson and King that asserted that human-chimpanzee divergence is about 1%. Humans share a common ancestor with chimpanzees, and the rapid evolution of chimpanzees and humans, along with gorillas and bonobos, has led to difficulties in creating an accurate lineage or tree topology. Chimpanzees and humans were found to be a monophyletic clade, leading to the question of how closely related the two are.