
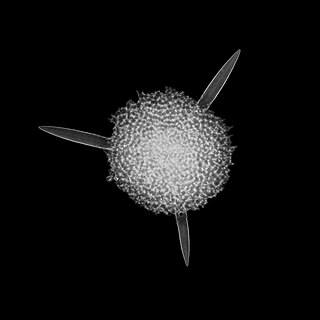
The Acantharea (Acantharia) are a group of radiolarian protozoa, distinguished mainly by their strontium sulfate skeletons. Acantharians are heterotrophic marine microplankton that range in size from about 200 microns in diameter up to several millimeters. Some acantharians have photosynthetic endosymbionts and hence are considered mixotrophs.

The Radiolaria, also called Radiozoa, are protozoa of diameter 0.1–0.2 mm that produce intricate mineral skeletons, typically with a central capsule dividing the cell into the inner and outer portions of endoplasm and ectoplasm. The elaborate mineral skeleton is usually made of silica. They are found as zooplankton throughout the global ocean. As zooplankton, radiolarians are primarily heterotrophic, but many have photosynthetic endosymbionts and are, therefore, considered mixotrophs. The skeletal remains of some types of radiolarians make up a large part of the cover of the ocean floor as siliceous ooze. Due to their rapid change as species and intricate skeletons, radiolarians represent an important diagnostic fossil found from the Cambrian onwards.

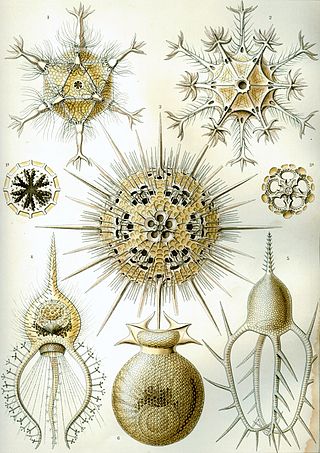
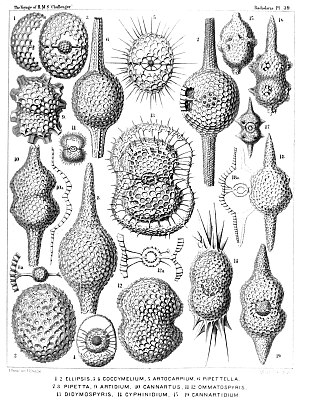
The polycystines are a group of radiolarians. They include the vast majority of the fossil radiolaria, as their skeletons are abundant in marine sediments, making them one of the most common groups of microfossils. These skeletons are composed of opaline silica. In some it takes the form of relatively simple spicules, but in others it forms more elaborate lattices, such as concentric spheres with radial spines or sequences of conical chambers. Two of the orders belonging to this group are the radially-symmetrical Spumellaria, dating back to the late Cambrian period, and the bilaterally-symmetrical Nasselaria, whose origin is placed within the lower Devonian.
Phaeodarea, or Phaeodaria, is a group of amoeboid cercozoan organisms. They are traditionally considered radiolarians, but in molecular trees do not appear to be close relatives of the other groups, and are instead placed among the Cercozoa. They are distinguished by the structure of their central capsule and by the presence of a phaeodium, an aggregate of waste particles within the cell.

A microfossil is a fossil that is generally between 0.001 mm and 1 mm in size, the visual study of which requires the use of light or electron microscopy. A fossil which can be studied with the naked eye or low-powered magnification, such as a hand lens, is referred to as a macrofossil.

Demosponges (Demospongiae) are the most diverse class in the phylum Porifera. They include 76.2% of all species of sponges with nearly 8,800 species worldwide. They are sponges with a soft body that covers a hard, often massive skeleton made of calcium carbonate, either aragonite or calcite. They are predominantly leuconoid in structure. Their "skeletons" are made of spicules consisting of fibers of the protein spongin, the mineral silica, or both. Where spicules of silica are present, they have a different shape from those in the otherwise similar glass sponges. Some species, in particular from the Antarctic, obtain the silica for spicule building from the ingestion of siliceous diatoms.

Spumellaria is an order of radiolarians in the class Polycystinea. They are ameboid protists appearing in abundance in the world's oceans, possessing a radially-symmetrical silica (opal) skeleton that has ensured their preservation in fossil records. They belong among the oldest Polycystine organisms, dating back to the lower Cambrian. Historically, many concentric radiolarians have been included in the Spumellaria order based on the absence of the initial spicular system, an early-develop structure that, by its lacking, sets them apart from Entactinaria despite their similar morphology. Living exemplars of the order feed by catching prey, such as copepod nauplii or tintinnids, on the adhesive ends of their pseudopodia extending radially from their skeleton; however, some have been observed as mixotrophs living in symbiosis with various photosynthetic algal organisms such as dinoflagellates, cyanobacteria, prasinophytes or haptophytes, which may cause their distribution to center in the greatest abundance and diversity within trophical waters.
Spongodiscidae is a family of radiolarians in the order Spumellaria. According to the original description by Ernst Haeckel, members of the family have a flat discoidal shell, in which a simple spherical central chamber is surrounded by an irregular spongy framework.

Stylosphaeridae is a family of radiolarians in the order Spumellaria. According to the original description by Ernst Haeckel, members of the family have a spherical central capsule within a fenestrated spherical siliceous shell, with two radial spines opposite in one axis. They are solitary. i.e. not associated in colonies.
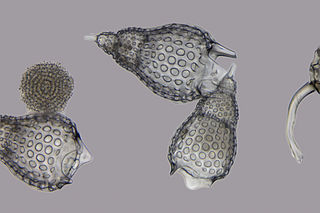
Nassellaria is an order of Rhizaria belonging to the class Radiolaria. The organisms of this order are characterized by a skeleton cross link with a cone or ring.
Cenellipsis is a genus of radiolarians in the order Spumellaria. The genus is extant but there are also fossil species.

Phaeocystida, also known as Phaeocystina, is a group of cercozoans in the class Phaeodarea. It was first described by Ernst Haeckel in 1887 and treated traditionally as a suborder, but later was raised to order level until Cavalier-Smith's classification lowered it again to suborder level. It belongs to the order Eodarida, characterised by simpler silica skeletons or a lack thereof.
Cornutanna is a genus of radiolarians in the order Nassellaria.
The genus Stylodictya belongs to a group of organisms called the Radiolaria. Radiolarians are amoeboid protists found as zooplankton in oceans around the world and are typically identified by their ornate skeletons.

Collozoum is a radiolarian genus formerly reported in the subfamily Sphaerozoidae, now reported descending from the order Collodaria. The genus contains bioluminescent species. It is a genus of colonial radiolarians.

Acrosphaera is a radiolarian genus in the Collosphaeridae. The genus contains bioluminescent species. It is a genus of colonial radiolarians.
Myxosphaera is a radiolarian genus in the Collosphaeridae. The genus may contain bioluminescent species. It is a genus of colonial radiolarians.

Many protists have protective shells or tests, usually made from silica (glass) or calcium carbonate (chalk). Protists are mostly single-celled and microscopic. Their shells are often tough, mineralised forms that resist degradation, and can survive the death of the protist as a microfossil. Although protists are typically very small, they are ubiquitous. Their numbers are such that their shells play a huge part in the formation of ocean sediments and in the global cycling of elements and nutrients.
Vitalia Viktorovna Reshetnyak (Решетняк) (1925–2015) was a Soviet protozoologist and marine biologist, specialising in Radiolaria, Phaeodorea and Acantharea.
A protist is any eukaryotic organism that is not an animal, plant, or fungus. While it is likely that protists share a common ancestor, the last eukaryotic common ancestor, the exclusion of other eukaryotes means that protists do not form a natural group, or clade. Therefore, some protists may be more closely related to animals, plants, or fungi than they are to other protists. However, like algae, invertebrates and protozoans, the grouping is used for convenience.
