The ovary is a gonad in the female reproductive system that produces ova. When an ovum is released, this travels through the fallopian tube into the uterus. There is an ovary found on the left and the right side of the body. The ovaries also secrete hormones that play a role in the menstrual cycle and fertility. The ovary progresses through many stages beginning in the prenatal period through menopause. It is also an endocrine gland because of the various hormones that it secretes.

The menstrual cycle is a series of natural changes in hormone production and the structures of the uterus and ovaries of the female reproductive system that makes pregnancy possible. The ovarian cycle controls the production and release of eggs and the cyclic release of estrogen and progesterone. The uterine cycle governs the preparation and maintenance of the lining of the uterus (womb) to receive an embryo. These cycles are concurrent and coordinated, normally last between 21 and 35 days, with a median length of 28 days, and continue for about 30–45 years.

Ovulation is the release of eggs from the ovaries. In women, this event occurs when the ovarian follicles rupture and release the secondary oocyte ovarian cells. After ovulation, during the luteal phase, the egg will be available to be fertilized by sperm. In addition, the uterine lining (endometrium) is thickened to be able to receive a fertilized egg. If no conception occurs, the uterine lining as well as the egg will be shed during menstruation.
Luteinizing hormone is a hormone produced by gonadotropic cells in the anterior pituitary gland. The production of LH is regulated by gonadotropin-releasing hormone (GnRH) from the hypothalamus. In females, an acute rise of LH known as an LH surge, triggers ovulation and development of the corpus luteum. In males, where LH had also been called interstitial cell–stimulating hormone (ICSH), it stimulates Leydig cell production of testosterone. It acts synergistically with follicle-stimulating hormone (FSH).

Follicle-stimulating hormone (FSH) is a gonadotropin, a glycoprotein polypeptide hormone. FSH is synthesized and secreted by the gonadotropic cells of the anterior pituitary gland and regulates the development, growth, pubertal maturation, and reproductive processes of the body. FSH and luteinizing hormone (LH) work together in the reproductive system.
Israel Hanukoglu is a Turkish-born Israeli scientist. He is a full professor of biochemistry and molecular biology at Ariel University and former science and technology adviser to the prime minister of Israel (1996–1999). He is founder of Israel Science and Technology Directory.

An ovarian follicle is a roughly spheroid cellular aggregation set found in the ovaries. It secretes hormones that influence stages of the menstrual cycle. At the time of puberty, women have approximately 200,000 to 300,000 follicles, each with the potential to release an egg cell (ovum) at ovulation for fertilization. These eggs are developed once every menstrual cycle with around 450–500 being ovulated during a woman's reproductive lifetime.

The corpus albicans is the regressed form of the corpus luteum. As the corpus luteum is being broken down by macrophages, fibroblasts lay down type I collagen, forming the corpus albicans. This process is called "luteolysis". The remains of the corpus albicans may persist as a scar on the surface of the ovary.

A granulosa cell or follicular cell is a somatic cell of the sex cord that is closely associated with the developing female gamete in the ovary of mammals.
The estrous cycle is a set of recurring physiological changes induced by reproductive hormones in females of mammalian subclass Theria. Estrous cycles start after sexual maturity in females and are interrupted by anestrous phases, otherwise known as "rest" phases, or by pregnancies. Typically, estrous cycles repeat until death. These cycles are widely variable in duration and frequency depending on the species. Some animals may display bloody vaginal discharge, often mistaken for menstruation. Many mammals used in commercial agriculture, such as cattle and sheep, may have their estrous cycles artificially controlled with hormonal medications for optimum productivity. The male equivalent, seen primarily in ruminants, is called rut.

In biology, folliculogenesis is the maturation of the ovarian follicle, a densely packed shell of somatic cells that contains an immature oocyte. Folliculogenesis describes the progression of a number of small primordial follicles into large preovulatory follicles that occurs in part during the menstrual cycle.
Luteolysis is the structural and functional degradation of the corpus luteum, which occurs at the end of the luteal phase of both the estrous and menstrual cycles in the absence of pregnancy.
The menstrual cycle is on average 28 days in length. It begins with menses during the follicular phase, followed by ovulation and ending with the luteal phase. Unlike the follicular phase which can vary in length among individuals, the luteal phase is typically fixed at approximately 14 days and is characterized by changes to hormone levels, such as an increase in progesterone and estrogen levels, decrease in gonadotropins such as follicle-stimulating hormone (FSH) and luteinizing hormone (LH), changes to the endometrial lining to promote implantation of the fertilized egg, and development of the corpus luteum. In the absence of fertilization by sperm, the corpus luteum atrophies leading to a decrease in progesterone and estrogen, an increase in FSH and LH, and shedding of the endometrial lining (menses) to begin the menstrual cycle again.
The follicular phase, also known as the preovulatory phase or proliferative phase, is the phase of the estrous cycle during which follicles in the ovary mature from primary follicle to a fully mature graafian follicle. It ends with ovulation. The main hormones controlling this stage are secretion of gonadotropin-releasing hormones, which are follicle-stimulating hormones and luteinising hormones. They are released by pulsatile secretion. The duration of the follicular phase can differ depending on the length of the menstrual cycle, while the luteal phase is usually stable, does not really change and lasts 14 days.
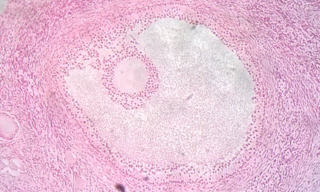
Follicular atresia refers to the process in which a follicle fails to develop, thus preventing it from ovulating and releasing an egg. It is a normal, naturally occurring progression that occurs as mammalian ovaries age. Approximately 1% of mammalian follicles in ovaries undergo ovulation and the remaining 99% of follicles go through follicular atresia as they cycle through the growth phases. In summary, follicular atresia is a process that leads to the follicular loss and loss of oocytes, and any disturbance or loss of functionality of this process can lead to many other conditions.
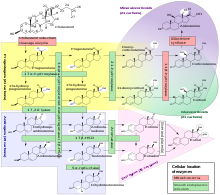
Theca interna cells express receptors for luteinizing hormone (LH) to produce androstenedione, which via a few steps, gives the granulosa the precursor for estrogen manufacturing.

Cholesterol side-chain cleavage enzyme is commonly referred to as P450scc, where "scc" is an acronym for side-chain cleavage. P450scc is a mitochondrial enzyme that catalyzes conversion of cholesterol to pregnenolone. This is the first reaction in the process of steroidogenesis in all mammalian tissues that specialize in the production of various steroid hormones.
The theca folliculi comprise a layer of the ovarian follicles. They appear as the follicles become secondary follicles.
Theca lutein cyst is a type of bilateral functional ovarian cyst filled with clear, straw-colored fluid. These cysts result from exaggerated physiological stimulation due to elevated levels of beta-human chorionic gonadotropin (beta-hCG) or hypersensitivity to beta-hCG. On ultrasound and MRI, theca lutein cysts appear in multiples on ovaries that are enlarged.
Gonadotropin surge-attenuating factor (GnSAF) is a nonsteroidal ovarian hormone produced by the granulosa cells of small antral ovarian follicles in females. GnSAF is involved in regulating the secretion of luteinizing hormone (LH) from the anterior pituitary and the ovarian cycle. During the early to mid-follicular phase of the ovarian cycle, GnSAF acts on the anterior pituitary to attenuate LH release, limiting the secretion of LH to only basal levels. At the transition between follicular and luteal phase, GnSAF bioactivity declines sufficiently to permit LH secretion above basal levels, resulting in the mid-cycle LH surge that initiates ovulation. In normally ovulating women, the LH surge only occurs when the oocyte is mature and ready for extrusion. GnSAF bioactivity is responsible for the synchronised, biphasic nature of LH secretion.