
Basidiomycota is one of two large divisions that, together with the Ascomycota, constitute the subkingdom Dikarya within the kingdom Fungi. Members are known as basidiomycetes. More specifically, Basidiomycota includes these groups: agarics, puffballs, stinkhorns, bracket fungi, other polypores, jelly fungi, boletes, chanterelles, earth stars, smuts, bunts, rusts, mirror yeasts, and Cryptococcus, the human pathogenic yeast.

Ascomycota is a phylum of the kingdom Fungi that, together with the Basidiomycota, forms the subkingdom Dikarya. Its members are commonly known as the sac fungi or ascomycetes. It is the largest phylum of Fungi, with over 64,000 species. The defining feature of this fungal group is the "ascus", a microscopic sexual structure in which nonmotile spores, called ascospores, are formed. However, some species of Ascomycota are asexual and thus do not form asci or ascospores. Familiar examples of sac fungi include morels, truffles, brewers' and bakers' yeast, dead man's fingers, and cup fungi. The fungal symbionts in the majority of lichens such as Cladonia belong to the Ascomycota.

The smuts are multicellular fungi characterized by their large numbers of teliospores. The smuts get their name from a Germanic word for 'dirt' because of their dark, thick-walled, and dust-like teliospores. They are mostly Ustilaginomycetes and comprise seven of the 15 orders of the subphylum. Most described smuts belong to two orders, Ustilaginales and Tilletiales. The smuts are normally grouped with the other basidiomycetes because of their commonalities concerning sexual reproduction.

Fungi are a diverse group of organisms that employ a huge variety of reproductive strategies, ranging from fully asexual to almost exclusively sexual species. Most species can reproduce both sexually and asexually, alternating between haploid and diploid forms. This contrasts with most multicellular eukaryotes such as mammals, where the adults are usually diploid and produce haploid gametes which combine to form the next generation. In fungi, both haploid and diploid forms can reproduce – haploid individuals can undergo asexual reproduction while diploid forms can produce gametes that combine to give rise to the next generation.

The Ustilaginales are an order of fungi within the class Ustilaginomycetes. The order contained 8 families, 49 genera, and 851 species in 2008.

The Ustilaginomycotina is a subdivision within the division Basidiomycota of the kingdom Fungi. It consists of the classes Ustilaginomycetes and Exobasidiomycetes, and in 2014 the subdivision was reclassified and the two additional classes Malasseziomycetes and Monilielliomycetes added. The name was first published by Doweld in 2001; Bauer and colleagues later published it in 2006 as an isonym. Ustilagomycotina and Agaricomycotina are considered to be sister groups, and they are in turn sister groups to the subdivision Pucciniomycotina.

Exobasidium vaccinii, commonly known as “red leaf disease,” or “Azalea Gall,” is a biotrophic species of fungus that causes galls on ericaceous plant species, such as blueberry and azalea. Exobasidium vaccinii is considered the type species of the Exobasidium genus. As a member of the Ustilagomycota, it is a basidiomycete closely related to smut fungi. Karl Wilhelm Gottlieb Leopold Fuckel first described the species in 1861 under the basionym Fusidium vaccinii, but in 1867 Mikhail Stepanovich Voronin later placed it in the genus Exobasidium. The type specimen is from Germany, and it is held in the Swedish Museum of Natural History. Exobasidium vaccinii, in current definition from John Axel Nannfeldt in 1981, is limited on the host Vaccinium vitis-idaea. This idea is used in most recent papers on E. vaccinii.

Pucciniomycotina is a subdivision of fungus within the division Basidiomycota. The subdivision contains 10 classes, 21 orders, and 38 families. Over 8400 species of Pucciniomycotina have been described - more than 8% of all described fungi. The subdivision is considered a sister group to Ustilaginomycotina and Agaricomycotina, which may share the basal lineage of Basidiomycota, although this is uncertain due to low support for placement between the three groups. The group was known as Urediniomycetes until 2006, when it was elevated from a class to a subdivision and named after the largest order in the group, Pucciniales.
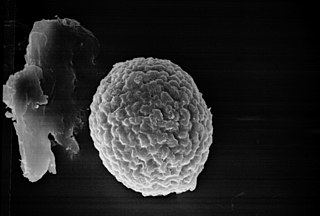
The Tilletiales are an order of smut fungi in the class Exobasidiomycetes. It is a monotypic order, consisting of a single family, the Tilletiaceae, which contains seven genera. The roughly 150 species in the Tilletiales all infect hosts of the grass family, except for species of Erratomyces, which occur on legumes.

The Mixiomycetes are a class of fungi in the Pucciniomycotina subdivision of the Basidiomycota. The class contains a single order, the Mixiales, which in turn contains a single family, the Mixiaceae that circumscribes the monotypic genus Mixia. Only one species has been described to date, Mixia osmundae; this species was originally named Taphrina osmundae by Japanese mycologist Toji Nishida in 1911. It is characterized by having multinucleate hyphae, and by producing multiple spores on sporogenous cells.

Pucciniomycetes is a diverse class of fungi in the subphylum Pucciniomycotina of phylum Basidiomycota. The class contains 5 orders, 21 families, 190 genera, and approximately 8,016 species. It has been estimated that this class contains about one third of all teleomorphic basidiomycetes. Pucciniomycetes contains many economically important plant pathogenic fungal rusts; the order Pucciniales is the largest clade in this class, representing approximately 7,000 species.

Helicobasidium is a genus of fungi in the subdivision Pucciniomycotina. Basidiocarps are corticioid (patch-forming) and are typically violet to purple. Microscopically they have auricularioid basidia. Asexual anamorphs, formerly referred to the genus Thanatophytum, produce sclerotia. Conidia-bearing anamorphs are parasitic on rust fungi and are currently still referred to the genus Tuberculina.
Robert Bauer was a German mycologist, specialising in rust (Uredinales) and smut (Ustilaginomycetes) fungi.

The Urocystidales are an order of fungi within the class Ustilaginomycetes. The order contains 6 families and about 400 genera. They are a sister order to Ustilaginales.

The Doassansiaceae are a family of fungi in the division Basidiomycota and order of Doassansiales. The family contains 11 genera and about 58 species. They have a widespread distribution. Doassansiaceae is also known and classified as a smut fungi.
The Melaniellaceae are a family of fungi in the division Basidiomycota and order of Doassansiales. The family contains 1 genera and 2 species. They have a distribution in south and south-east Asia.

The Rhamphosporaceae is a family of fungi in the division Basidiomycota and order of Doassansiales. The monotypic family only contains 1 genus; RhamphosporaD.D.Cunn. and just 1 species, Rhamphospora nymphaeaeD.D.Cunn. It is found on the leaves of waterlilies causing spots.

The Platygloeales are an order of fungi in the class Pucciniomycetes. Species in the order have auricularioid basidia and are typically plant parasites on mosses, ferns, and angiosperms, though Platygloea species appear to be saprotrophic.
Naohidea sebacea is a species of fungus in the order Naohideales. The order is currently monotypic, having only one family, one genus, and one species. Basidiocarps of Naohidea sebacea form small, gelatinous pustules on wood-inhabiting species of Botryosphaeriaceae. Microscopically, they produce long, slender, auricularioid basidia and amygdaliform (almond-shaped) basidiospores.
Insolibasidium deformans is a species of fungus belonging to the order Platygloeales. It is currently the only species in the monotypic genus Insolibasidium. The fungus parasitizes leaves of various Lonicera species, causing honeysuckle leaf blight, a commercially significant disease in plant nurseries.


