
Hydrogen is a chemical element; it has symbol H and atomic number 1. It is the lightest element and, at standard conditions, is a gas of diatomic molecules with the formula H2. It is colorless, odorless, tasteless, non-toxic, and highly combustible. Hydrogen is the most abundant chemical substance in the universe, constituting roughly 75% of all normal matter. Stars such as the Sun are mainly composed of hydrogen in the plasma state. Most of the hydrogen on Earth exists in molecular forms such as water and organic compounds. For the most common isotope of hydrogen each atom has one proton, one electron, and no neutrons.

The Miller–Urey experiment (or Miller experiment) was an experiment in chemical synthesis carried out in 1952 that simulated the conditions thought at the time to be present in the atmosphere of the early, prebiotic Earth. It is seen as one of the first successful experiments demonstrating the synthesis of organic compounds from inorganic constituents in an origin of life scenario. The experiment used methane (CH4), ammonia (NH3), hydrogen (H2), in ratio 2:2:1, and water (H2O). Applying an electric arc (the latter simulating lightning) resulted in the production of amino acids.

A heterotroph is an organism that cannot produce its own food, instead taking nutrition from other sources of organic carbon, mainly plant or animal matter. In the food chain, heterotrophs are primary, secondary and tertiary consumers, but not producers. Living organisms that are heterotrophic include all animals and fungi, some bacteria and protists, and many parasitic plants. The term heterotroph arose in microbiology in 1946 as part of a classification of microorganisms based on their type of nutrition. The term is now used in many fields, such as ecology, in describing the food chain.

In biochemistry, chemosynthesis is the biological conversion of one or more carbon-containing molecules and nutrients into organic matter using the oxidation of inorganic compounds or ferrous ions as a source of energy, rather than sunlight, as in photosynthesis. Chemoautotrophs, organisms that obtain carbon from carbon dioxide through chemosynthesis, are phylogenetically diverse. Groups that include conspicuous or biogeochemically important taxa include the sulfur-oxidizing Gammaproteobacteria, the Campylobacterota, the Aquificota, the methanogenic archaea, and the neutrophilic iron-oxidizing bacteria.
Methanogens are microorganisms that produce methane as a metabolic byproduct in hypoxic conditions. They belong to the domain Archaea and are members of the phylum Euryarchaeota. Methanogens are common in wetlands, where they are responsible for marsh gas, and can occur in the digestive tracts of animals including ruminants and humans, where they are responsible for the methane content of belching and flatulence. In marine sediments, the biological production of methane, termed methanogenesis, is generally confined to where sulfates are depleted below the top layers and methanogens play an indispensable role in anaerobic wastewater treatments. Other methanogens are extremophiles, found in environments such as hot springs and submarine hydrothermal vents as well as in the "solid" rock of Earth's crust, kilometers below the surface.
Methanogenesis or biomethanation is the formation of methane coupled to energy conservation by microbes known as methanogens. Organisms capable of producing methane for energy conservation have been identified only from the domain Archaea, a group phylogenetically distinct from both eukaryotes and bacteria, although many live in close association with anaerobic bacteria. The production of methane is an important and widespread form of microbial metabolism. In anoxic environments, it is the final step in the decomposition of biomass. Methanogenesis is responsible for significant amounts of natural gas accumulations, the remainder being thermogenic.
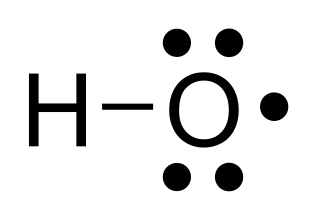
The hydroxyl radical, •HO, is the neutral form of the hydroxide ion (HO–). Hydroxyl radicals are highly reactive and consequently short-lived; however, they form an important part of radical chemistry. Most notably hydroxyl radicals are produced from the decomposition of hydroperoxides (ROOH) or, in atmospheric chemistry, by the reaction of excited atomic oxygen with water. It is also an important radical formed in radiation chemistry, since it leads to the formation of hydrogen peroxide and oxygen, which can enhance corrosion and SCC in coolant systems subjected to radioactive environments. Hydroxyl radicals are also produced during UV-light dissociation of H2O2 (suggested in 1879) and likely in Fenton chemistry, where trace amounts of reduced transition metals catalyze peroxide-mediated oxidations of organic compounds.

Sulfate-reducing microorganisms (SRM) or sulfate-reducing prokaryotes (SRP) are a group composed of sulfate-reducing bacteria (SRB) and sulfate-reducing archaea (SRA), both of which can perform anaerobic respiration utilizing sulfate (SO2−
4) as terminal electron acceptor, reducing it to hydrogen sulfide (H2S). Therefore, these sulfidogenic microorganisms "breathe" sulfate rather than molecular oxygen (O2), which is the terminal electron acceptor reduced to water (H2O) in aerobic respiration.

Serpentinization is a hydration and metamorphic transformation of ferromagnesian minerals, such as olivine and pyroxene, in mafic and ultramafic rock to produce serpentinite. Minerals formed by serpentinization include the serpentine group minerals, brucite, talc, Ni-Fe alloys, and magnetite. The mineral alteration is particularly important at the sea floor at tectonic plate boundaries.
Microbial metabolism is the means by which a microbe obtains the energy and nutrients it needs to live and reproduce. Microbes use many different types of metabolic strategies and species can often be differentiated from each other based on metabolic characteristics. The specific metabolic properties of a microbe are the major factors in determining that microbe's ecological niche, and often allow for that microbe to be useful in industrial processes or responsible for biogeochemical cycles.
In biology, syntrophy, syntrophism, or cross-feeding is the cooperative interaction between at least two microbial species to degrade a single substrate. This type of biological interaction typically involves the transfer of one or more metabolic intermediates between two or more metabolically diverse microbial species living in close proximity to each other. Thus, syntrophy can be considered an obligatory interdependency and a mutualistic metabolism between different microbial species, wherein the growth of one partner depends on the nutrients, growth factors, or substrates provided by the other(s).

Biohydrogen is H2 that is produced biologically. Interest is high in this technology because H2 is a clean fuel and can be readily produced from certain kinds of biomass, including biological waste. Furthermore some photosynthetic microorganisms are capable to produce H2 directly from water splitting using light as energy source.
Hydrogen-oxidizing bacteria are a group of facultative autotrophs that can use hydrogen as an electron donor. They can be divided into aerobes and anaerobes. The former use hydrogen as an electron donor and oxygen as an acceptor while the latter use sulphate or nitrogen dioxide as electron acceptors. Species of both types have been isolated from a variety of environments, including fresh waters, sediments, soils, activated sludge, hot springs, hydrothermal vents and percolating water.

Methane is a chemical compound with the chemical formula CH4. It is a group-14 hydride, the simplest alkane, and the main constituent of natural gas. The relative abundance of methane on Earth makes it an economically attractive fuel, although capturing and storing it poses technical challenges due to its gaseous state under normal conditions for temperature and pressure.
An autotroph is an organism that produces complex organic compounds using carbon from simple substances such as carbon dioxide, generally using energy from light (photosynthesis) or inorganic chemical reactions (chemosynthesis). They convert an abiotic source of energy into energy stored in organic compounds, which can be used by other organisms. Autotrophs do not need a living source of carbon or energy and are the producers in a food chain, such as plants on land or algae in water. Autotrophs can reduce carbon dioxide to make organic compounds for biosynthesis and as stored chemical fuel. Most autotrophs use water as the reducing agent, but some can use other hydrogen compounds such as hydrogen sulfide.
Sulfurimonas is a bacterial genus within the class of Campylobacterota, known for reducing nitrate, oxidizing both sulfur and hydrogen, and containing Group IV hydrogenases. This genus consists of four species: Sulfurimonas autorophica, Sulfurimonas denitrificans, Sulfurimonas gotlandica, and Sulfurimonas paralvinellae. The genus' name is derived from "sulfur" in Latin and "monas" from Greek, together meaning a “sulfur-oxidizing rod”. The size of the bacteria varies between about 1.5-2.5 μm in length and 0.5-1.0 μm in width. Members of the genus Sulfurimonas are found in a variety of different environments which include deep sea-vents, marine sediments, and terrestrial habitats. Their ability to survive in extreme conditions is attributed to multiple copies of one enzyme. Phylogenetic analysis suggests that members of the genus Sulfurimonas have limited dispersal ability and its speciation was affected by geographical isolation rather than hydrothermal composition. Deep ocean currents affect the dispersal of Sulfurimonas spp., influencing its speciation. As shown in the MLSA report of deep-sea hydrothermal vents Campylobacterota, Sulfurimonas has a higher dispersal capability compared with deep sea hydrothermal vent thermophiles, indicating allopatric speciation.

Enceladus Life Finder (ELF) is a proposed astrobiology mission concept for a NASA spacecraft intended to assess the habitability of the internal aquatic ocean of Enceladus, which is Saturn's sixth-largest moon of at least 146 total moons, and seemingly similar in chemical makeup to comets. The spaceprobe would orbit Saturn and fly through Enceladus's geyser-like plumes multiple times. It would be powered by energy supplied from solar panels on the spacecraft.

The hydrothermal vent microbial community includes all unicellular organisms that live and reproduce in a chemically distinct area around hydrothermal vents. These include organisms in the microbial mat, free floating cells, or bacteria in an endosymbiotic relationship with animals. Chemolithoautotrophic bacteria derive nutrients and energy from the geological activity at Hydrothermal vents to fix carbon into organic forms. Viruses are also a part of the hydrothermal vent microbial community and their influence on the microbial ecology in these ecosystems is a burgeoning field of research.
Formatotrophs are organisms that can assimilate formate or formic acid to use as a carbon source or for reducing power. Some authors classify formatotrophs as one of the five trophic groups of methanogens, which also include hydrogenotrophs, acetotrophs, methylotrophs, and alcoholotrophs. Formatotrophs have garnered attention for applications in biotechnology as part of a "formate bioeconomy" in which synthesized formate could be used as a nutrient for microoganisms. Formate can be electrochemically synthesized from CO2 and renewable energy, and formatotrophs may be genetically modified to enhance production of biochemical products to be used as biofuels. Technical limitations in culturing formatotrophs have limited the discovery of natural formatotrophs and impeded research on their formate-metabolizing enzymes, which are of interest for applications in carbon sequestration and astrobiology.

The prebiotic atmosphere is the second atmosphere present on Earth before today's biotic, oxygen-rich third atmosphere, and after the first atmosphere of Earth's formation. The formation of the Earth, roughly 4.5 billion years ago, involved multiple collisions and coalescence of planetary embryos. This was followed by a <100 million year period on Earth where a magma ocean was present, the atmosphere was mainly steam, and surface temperatures reached up to 8,000 K (14,000 °F). Earth's surface then cooled and the atmosphere stabilized, establishing the prebiotic atmosphere. The environmental conditions during this time period were quite different from today: the Sun was ~30% dimmer overall yet brighter at ultraviolet and x-ray wavelengths, there was a liquid ocean, it is unknown if there were continents but oceanic islands were likely, Earth's interior chemistry was different, and there was a larger flux of impactors hitting Earth's surface.
