
A diatom is any member of a large group comprising several genera of algae, specifically microalgae, found in the oceans, waterways and soils of the world. Living diatoms make up a significant portion of the Earth's biomass: they generate about 20 to 50 percent of the oxygen produced on the planet each year, take in over 6.7 billion tonnes of silicon each year from the waters in which they live, and constitute nearly half of the organic material found in the oceans. The shells of dead diatoms can reach as much as a half-mile deep on the ocean floor, and the entire Amazon basin is fertilized annually by 27 million tons of diatom shell dust transported by transatlantic winds from the African Sahara, much of it from the Bodélé Depression, which was once made up of a system of fresh-water lakes.

Chert is a hard, fine-grained sedimentary rock composed of microcrystalline or cryptocrystalline quartz, the mineral form of silicon dioxide (SiO2). Chert is characteristically of biological origin, but may also occur inorganically as a chemical precipitate or a diagenetic replacement, as in petrified wood.

The Radiolaria, also called Radiozoa, are protozoa of diameter 0.1–0.2 mm that produce intricate mineral skeletons, typically with a central capsule dividing the cell into the inner and outer portions of endoplasm and ectoplasm. The elaborate mineral skeleton is usually made of silica. They are found as zooplankton throughout the global ocean. As zooplankton, radiolarians are primarily heterotrophic, but many have photosynthetic endosymbionts and are, therefore, considered mixotrophs. The skeletal remains of some types of radiolarians make up a large part of the cover of the ocean floor as siliceous ooze. Due to their rapid change as species and intricate skeletons, radiolarians represent an important diagnostic fossil found from the Cambrian onwards.

The biological pump (or ocean carbon biological pump or marine biological carbon pump) is the ocean's biologically driven sequestration of carbon from the atmosphere and land runoff to the ocean interior and seafloor sediments. In other words, it is a biologically mediated process which results in the sequestering of carbon in the deep ocean away from the atmosphere and the land. The biological pump is the biological component of the "marine carbon pump" which contains both a physical and biological component. It is the part of the broader oceanic carbon cycle responsible for the cycling of organic matter formed mainly by phytoplankton during photosynthesis (soft-tissue pump), as well as the cycling of calcium carbonate (CaCO3) formed into shells by certain organisms such as plankton and mollusks (carbonate pump).

Diatomaceous earth, diatomite, celite or kieselgur/kieselguhr is a naturally occurring, soft, siliceous sedimentary rock that can be crumbled into a fine white to off-white powder. It has a particle size ranging from more than 3 mm to less than 1 μm, but typically 10 to 200 μm. Depending on the granularity, this powder can have an abrasive feel, similar to pumice powder, and has a low density as a result of its high porosity. The typical chemical composition of oven-dried diatomaceous earth is 80–90% silica, with 2–4% alumina, and 0.5–2% iron oxide.

A microfossil is a fossil that is generally between 0.001 mm and 1 mm in size, the visual study of which requires the use of light or electron microscopy. A fossil which can be studied with the naked eye or low-powered magnification, such as a hand lens, is referred to as a macrofossil.
High-nutrient, low-chlorophyll (HNLC) regions are regions of the ocean where the abundance of phytoplankton is low and fairly constant despite the availability of macronutrients. Phytoplankton rely on a suite of nutrients for cellular function. Macronutrients are generally available in higher quantities in surface ocean waters, and are the typical components of common garden fertilizers. Micronutrients are generally available in lower quantities and include trace metals. Macronutrients are typically available in millimolar concentrations, while micronutrients are generally available in micro- to nanomolar concentrations. In general, nitrogen tends to be a limiting ocean nutrient, but in HNLC regions it is never significantly depleted. Instead, these regions tend to be limited by low concentrations of metabolizable iron. Iron is a critical phytoplankton micronutrient necessary for enzyme catalysis and electron transport.
The carbonate compensation depth (CCD) is the depth, in the oceans, at which the rate of supply of calcium carbonates matches the rate of solvation. That is, solvation 'compensates' supply. Below the CCD solvation is faster, so that carbonate particles dissolve and the carbonate shells (tests) of animals are not preserved. Carbonate particles cannot accumulate in the sediments where the sea floor is below this depth.

Pelagic sediment or pelagite is a fine-grained sediment that accumulates as the result of the settling of particles to the floor of the open ocean, far from land. These particles consist primarily of either the microscopic, calcareous or siliceous shells of phytoplankton or zooplankton; clay-size siliciclastic sediment; or some mixture of these. Trace amounts of meteoric dust and variable amounts of volcanic ash also occur within pelagic sediments. Based upon the composition of the ooze, there are three main types of pelagic sediments: siliceous oozes, calcareous oozes, and red clays.
Lithogenic silica (LSi) is silica (SiO2) derived from terrigenous rock (Igneous, metamorphic, and sedimentary), lithogenic sediments composed of the detritus of pre-existing rock, volcanic ejecta, extraterrestrial material, and minerals such silicate. Silica is the most abundant compound in the Earth's crust (59%) and the main component of almost every rock (>95%).
Biogenic silica (bSi), also referred to as opal, biogenic opal, or amorphous opaline silica, forms one of the most widespread biogenic minerals. For example, microscopic particles of silica called phytoliths can be found in grasses and other plants.

Radiolarite is a siliceous, comparatively hard, fine-grained, chert-like, and homogeneous sedimentary rock that is composed predominantly of the microscopic remains of radiolarians. This term is also used for indurated radiolarian oozes and sometimes as a synonym of radiolarian earth. However, radiolarian earth is typically regarded by Earth scientists to be the unconsolidated equivalent of a radiolarite. A radiolarian chert is well-bedded, microcrystalline radiolarite that has a well-developed siliceous cement or groundmass.

Marine sediment, or ocean sediment, or seafloor sediment, are deposits of insoluble particles that have accumulated on the seafloor. These particles have their origins in soil and rocks and have been transported from the land to the sea, mainly by rivers but also by dust carried by wind and by the flow of glaciers into the sea. Additional deposits come from marine organisms and chemical precipitation in seawater, as well as from underwater volcanoes and meteorite debris.
Hemipelagic sediment, or hemipelagite, is a type of marine sediment that consists of clay and silt-sized grains that are terrigenous and some biogenic material derived from the landmass nearest the deposits or from organisms living in the water. Hemipelagic sediments are deposited on continental shelves and continental rises, and differ from pelagic sediment compositionally. Pelagic sediment is composed of primarily biogenic material from organisms living in the water column or on the seafloor and contains little to no terrigenous material. Terrigenous material includes minerals from the lithosphere like feldspar or quartz. Volcanism on land, wind blown sediments as well as particulates discharged from rivers can contribute to Hemipelagic deposits. These deposits can be used to qualify climatic changes and identify changes in sediment provenances.
Reverse weathering generally refers to the formation of a clay neoformation that utilizes cations and alkalinity in a process unrelated to the weathering of silicates. More specifically reverse weathering refers to the formation of authigenic clay minerals from the reaction of 1) biogenic silica with aqueous cations or cation bearing oxides or 2) cation poor precursor clays with dissolved cations or cation bearing oxides.
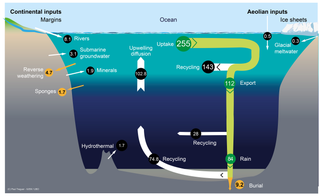
The silica cycle is the biogeochemical cycle in which biogenic silica is transported between the Earth's systems. Silicon is considered a bioessential element and is one of the most abundant elements on Earth. The silica cycle has significant overlap with the carbon cycle and plays an important role in the sequestration of carbon through continental weathering, biogenic export and burial as oozes on geologic timescales.

Many protists have protective shells or tests, usually made from silica (glass) or calcium carbonate (chalk). Protists are a diverse group of eukaryote organisms that are not plants, animals, or fungi. They are typically microscopic unicellular organisms that live in water or moist environments.

In geology, silicification is a petrification process in which silica-rich fluids seep into the voids of Earth materials, e.g., rocks, wood, bones, shells, and replace the original materials with silica (SiO2). Silica is a naturally existing and abundant compound found in organic and inorganic materials, including Earth's crust and mantle. There are a variety of silicification mechanisms. In silicification of wood, silica permeates into and occupies cracks and voids in wood such as vessels and cell walls. The original organic matter is retained throughout the process and will gradually decay through time. In the silicification of carbonates, silica replaces carbonates by the same volume. Replacement is accomplished through the dissolution of original rock minerals and the precipitation of silica. This leads to a removal of original materials out of the system. Depending on the structures and composition of the original rock, silica might replace only specific mineral components of the rock. Silicic acid (H4SiO4) in the silica-enriched fluids forms lenticular, nodular, fibrous, or aggregated quartz, opal, or chalcedony that grows within the rock. Silicification happens when rocks or organic materials are in contact with silica-rich surface water, buried under sediments and susceptible to groundwater flow, or buried under volcanic ashes. Silicification is often associated with hydrothermal processes. Temperature for silicification ranges in various conditions: in burial or surface water conditions, temperature for silicification can be around 25°−50°; whereas temperatures for siliceous fluid inclusions can be up to 150°−190°. Silicification could occur during a syn-depositional or a post-depositional stage, commonly along layers marking changes in sedimentation such as unconformities or bedding planes.
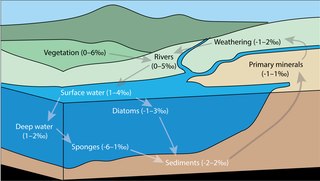
Silicon isotope biogeochemistry is the study of environmental processes using the relative abundance of Si isotopes. As the relative abundance of Si stable isotopes varies among different natural materials, the differences in abundance can be used to trace the source of Si, and to study biological, geological, and chemical processes. The study of stable isotope biogeochemistry of Si aims to quantify the different Si fluxes in the global biogeochemical silicon cycle, to understand the role of biogenic silica within the global Si cycle, and to investigate the applications and limitations of the sedimentary Si record as an environmental and palaeoceanographic proxy.
Biogenous ooze is marine sediment that accumulates on the seafloor and is a byproduct of the death and sink of the skeletal remains of marine organisms.







