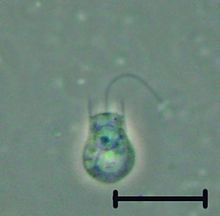
A flagellum is a hairlike appendage that protrudes from certain plant and animal sperm cells, from fungal spores (zoospores), and from a wide range of microorganisms to provide motility. Many protists with flagella are known as flagellates.
Choanocytes are cells that line the interior of asconoid, syconoid and leuconoid body types of sponges that contain a central flagellum, or cilium, surrounded by a collar of microvilli which are connected by a thin membrane.

The opisthokonts are a broad group of eukaryotes, including both the animal and fungus kingdoms. The opisthokonts, previously called the "Fungi/Metazoa group", are generally recognized as a clade. Opisthokonts together with Apusomonadida and Breviata comprise the larger clade Obazoa.

Sexual reproduction is an adaptive feature which is common to almost all multicellular organisms and various unicellular organisms. Currently, the adaptive advantage of sexual reproduction is widely regarded as a major unsolved problem in biology. As discussed below, one prominent theory is that sex evolved as an efficient mechanism for producing variation, and this had the advantage of enabling organisms to adapt to changing environments. Another prominent theory, also discussed below, is that a primary advantage of outcrossing sex is the masking of the expression of deleterious mutations. Additional theories concerning the adaptive advantage of sex are also discussed below. Sex does, however, come with a cost. In reproducing asexually, no time nor energy needs to be expended in choosing a mate and, if the environment has not changed, then there may be little reason for variation, as the organism may already be well-adapted. However, very few environments have not changed over the millions of years that reproduction has existed. Hence it is easy to imagine that being able to adapt to changing environment imparts a benefit. Sex also halves the amount of offspring a given population is able to produce. Sex, however, has evolved as the most prolific means of species branching into the tree of life. Diversification into the phylogenetic tree happens much more rapidly via sexual reproduction than it does by way of asexual reproduction.

Amorphea is a taxonomic supergroup that includes the basal Amoebozoa and Obazoa. That latter contains the Opisthokonta, which includes the Fungi, Animals and the Choanomonada, or Choanoflagellates. The taxonomic affinities of the members of this clade were originally described and proposed by Thomas Cavalier-Smith in 2002.

Proterospongia is a genus of single-celled aquatic organisms which form colonies. It belongs to the choanoflagellate class. As a colony-forming choanoflagellate, Proterospongia is of interest to scientists studying the mechanisms of intercellular signaling and adhesion present before animals appeared.
"Candidatus Midichloria" is a candidatus genus of Gram-negative, non-endospore-forming bacteria, with a bacillus shape around 0.45 µm in diameter and 1.2 µm in length. First described in 2004 with the temporary name IricES1, "Candidatus Midichloria" species are symbionts of several species of hard ticks. They live in the cells of the ovary of the females of this tick species. These bacteria have been observed in the mitochondria of the host cells, a trait that has never been described in any other symbiont of animals.
Metacaspases are members of the C14 class of cysteine proteases and thus related to caspases, orthocaspases and paracaspases. The metacaspases are arginine/lysine-specific, in contrast to caspases, which are aspartate-specific.

Holozoa is a clade of organisms that includes animals and their closest single-celled relatives, but excludes fungi and all other organisms. Together they amount to more than 1.5 million species of purely heterotrophic organisms, including around 300 unicellular species. It consists of various subgroups, namely Metazoa and the protists Choanoflagellata, Filasterea, Pluriformea and Ichthyosporea. Along with fungi and some other groups, Holozoa is part of the Opisthokonta, a supergroup of eukaryotes. Choanofila was previously used as the name for a group similar in composition to Holozoa, but its usage is discouraged now because it excludes animals and is therefore paraphyletic.

Guillardia is a genus of marine biflagellate cryptomonad algae with a plastid obtained through secondary endosymbiosis of a red alga.

The Filozoa are a monophyletic grouping within the Opisthokonta. They include animals and their nearest unicellular relatives.
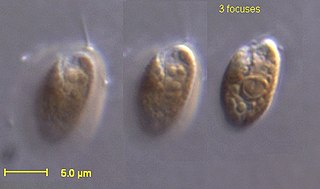
The eukaryotes constitute the domain of Eukarya or Eukaryota, organisms whose cells have a membrane-bound nucleus. All animals, plants, fungi, and many unicellular organisms are eukaryotes. They constitute a major group of life forms alongside the two groups of prokaryotes: the Bacteria and the Archaea. Eukaryotes represent a small minority of the number of organisms, but given their generally much larger size, their collective global biomass is much larger than that of prokaryotes.

Until the late 1950s, the Precambrian was not believed to have hosted multicellular organisms. However, with radiometric dating techniques, it has been found that fossils initially found in the Ediacara Hills in Southern Australia date back to the late Precambrian. These fossils are body impressions of organisms shaped like disks, fronds and some with ribbon patterns that were most likely tentacles.

Choanozoa is a clade of opisthokont eukaryotes consisting of the choanoflagellates (Choanoflagellatea) and the animals. The sister-group relationship between the choanoflagellates and animals has important implications for the origin of the animals. The clade was identified in 2015 by Graham Budd and Sören Jensen, who used the name Apoikozoa. The 2018 revision of the classification first proposed by the International Society of Protistologists in 2012 recommends the use of the name Choanozoa.

Salpingoeca rosetta is a species of Choanoflagellates in the family Salpingoecidae. It is a rare marine eukaryote consisting of a number of cells embedded in a jelly-like matrix. This organism demonstrates a very primitive level of cell differentiation and specialization. This is seen with flagellated cells and their collar structures that move the cell colony through the water.
Similar low level cellular differentiation and specification can also be seen in sponges. They also have collar cells and amoeboid cells arranged in a gelatinous matrix.
Unlike S. rosetta, sponges also have other cell-types that can perform different functions. Also, the collar cells of sponges beat within canals in the sponge body, whereas Salpingoeca rosetta's collar cells reside on the inside and it lacks internal canals. Despite these minor differences, there is strong evidence that Proterospongia and Metazoa are highly related.

Many protists have protective shells or tests, usually made from silica (glass) or calcium carbonate (chalk). Protists are a diverse group of eukaryote organisms that are not plants, animals, or fungi. They are typically microscopic unicellular organisms that live in water or moist environments.
Diaphanoeca grandis is a species of choanoflagellate in the family Acanthoecidae which is the type species of the genus Diaphanoeca. It is a unicellular micro-heterotroph with a large protective lorica that is found beneath sea ice in a wide distribution. The lorica is composed of silica and possibly originates from diatoms via Horizontal gene transfer.

Syssomonas is a monotypic genus of unicellular flagellated protists containing the species Syssomonas multiformis. It is a member of Pluriformea inside the lineage of Holozoa, a clade containing animals and their closest protistan relatives. It lives in freshwater habitats. It has a complex life cycle that includes unicellular amoeboid and flagellated phases, as well as multicellular aggregates, depending on the growth medium and nutritional state.
Crinolina isefiordensis is a species of choanoflagellate in the family Acanthoecidae. It is the type species of the genus Crinolina and is named for the first location of its collection, the Ise Fjord in Denmark.