
A chordate is a deuterostomic animal belonging to the phylum Chordata. All chordates possess, at some point during their larval or adult stages, five distinctive physical characteristics (synapomorphies) that distinguish them from other taxa. These five synapomorphies are a notochord, a hollow dorsal nerve cord, an endostyle or thyroid, pharyngeal slits, and a post-anal tail. The name "chordate" comes from the first of these synapomorphies, the notochord, which plays a significant role in chordate body plan structuring and movements. Chordates are also bilaterally symmetric, have a coelom, possess a closed circulatory system, and exhibit metameric segmentation.

Hemichordata is a phylum which consists of triploblastic, enterocoelomate, and bilaterally symmetrical marine deuterostome animals, generally considered the sister group of the echinoderms. They appear in the Lower or Middle Cambrian and include two main classes: Enteropneusta, and Pterobranchia. A third class, Planctosphaeroidea, is known only from the larva of a single species, Planctosphaera pelagica. The class Graptolithina, formerly considered extinct, is now placed within the pterobranchs, represented by a single living genus Rhabdopleura.

The Lophophorata or Tentaculata are a Lophotrochozoan clade consisting of the Brachiozoa and the Bryozoa. They have a lophophore. Molecular phylogenetic analyses suggest that lophophorates are protostomes, but on morphological grounds they have been assessed as deuterostomes. Fossil finds of the "tommotiid" Wufengella suggest that they evolved from worm-like animals that resembled annelids.

Vetulicolia is a phylum of bilaterian animals encompassing several extinct species belonging to the Cambrian period. The phylum was created by Degan Shu and his research team in 2001, and named after Vetulicola cuneata, the first species of the phylum described in 1987.

Bilateria is a large clade or infrakingdom of animals called bilaterians, characterized by bilateral symmetry during embryonic development. This means their body plans are laid around a longitudinal axis with a front and a rear end, as well as a left–right–symmetrical belly (ventral) and back (dorsal) surface. Nearly all bilaterians maintain a bilaterally symmetrical body as adults; the most notable exception is the echinoderms, which extend to pentaradial symmetry as adults, but are only bilaterally symmetrical as an embryo. Cephalization is also a characteristic feature among most bilaterians, where the special sense organs and central nerve ganglia become concentrated at the front/rostral end.

Amiskwia is a genus of soft-bodied animals known from fossils of the Middle Cambrian Lagerstätten both in the Burgess Shale in British Columbia, Canada and the Maotianshan shales of Yunnan Province, China. It is interpreted as a member of the clade Gnathifera sensu lato or as a stem-chaetognath.

Lophotrochozoa is a clade of protostome animals within the Spiralia. The taxon was established as a monophyletic group based on molecular evidence. The clade includes animals like annelids, molluscs, bryozoans, and brachiopods.
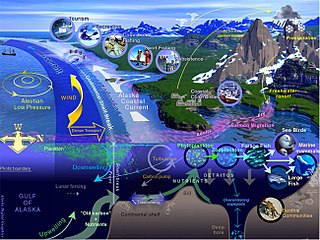
Marine life, sea life, or ocean life is the plants, animals, and other organisms that live in the salt water of seas or oceans, or the brackish water of coastal estuaries. At a fundamental level, marine life affects the nature of the planet. Marine organisms, mostly microorganisms, produce oxygen and sequester carbon. Marine life, in part, shape and protect shorelines, and some marine organisms even help create new land.

Animals are multicellular, eukaryotic organisms in the biological kingdom Animalia. With few exceptions, animals consume organic material, breathe oxygen, have myocytes and are able to move, can reproduce sexually, and grow from a hollow sphere of cells, the blastula, during embryonic development. Animals form a clade, meaning that they arose from a single common ancestor.

Deuterostomes are bilaterian animals of the superphylum Deuterostomia, typically characterized by their anus forming before the mouth during embryonic development. Deuterostomia is further divided into 4 phyla: Chordata, Echinodermata, Hemichordata, and the extinct Vetulicolia known from Cambrian fossils. The extinct clade Cambroernida is also thought to be a member of Deuterostomia.

Phoronids are a small phylum of marine animals that filter-feed with a lophophore, and build upright tubes of chitin to support and protect their soft bodies. They live in most of the oceans and seas, including the Arctic Ocean but excluding the Antarctic Ocean, and between the intertidal zone and about 400 meters down. Most adult phoronids are 2 cm long and about 1.5 mm wide, although the largest are 50 cm long.

Gnathifera is a clade of generally small spiralians characterized by complex jaws made of chitin. It comprises the phyla Gnathostomulida, Rotifera and Micrognathozoa. Chaetognatha has recently been recognised as closely related to the group, with it either being included within Gnathifera or the broader group Chaetognathifera. It may also include the Cycliophora.

Ooedigera peeli is an extinct vetulicolian from the Early Cambrian of North Greenland. The front body was flattened horizontally, oval-shaped, likely bearing a reticulated or anastomosing pattern, and had 5 evenly-spaced gill pouches along the midline. The tail was also bulbous and flattened horizontally, but was divided into 7 plates connected by flexible membranes, allowing movement. Ooedigera likely swam by moving side-to-side like a fish. It may have lived in an oxygen minimum zone alongside several predators in an ecosystem based on chemosynthetic microbial mats, and was possibly a deposit or filter feeder living near the seafloor.

Nephrozoa is a proposed major clade of bilaterian animals. It includes all bilaterians other than Xenacoelomorpha. It contrasts with the Xenambulacraria hypothesis, which instead posits that Xenacoelomorpha is most closely related to Ambulacraria. Which hypothesis is correct is controversial. Authors supporting the Xenambulacraria hypothesis have suggested that the genetic evidence used to support Nephrozoa may be due to systematic error.

Xenacoelomorpha is a small phylum of bilaterian invertebrate animals, consisting of two sister groups: xenoturbellids and acoelomorphs. This new phylum was named in February 2011 and suggested based on morphological synapomorphies, which was then confirmed by phylogenomic analyses of molecular data.

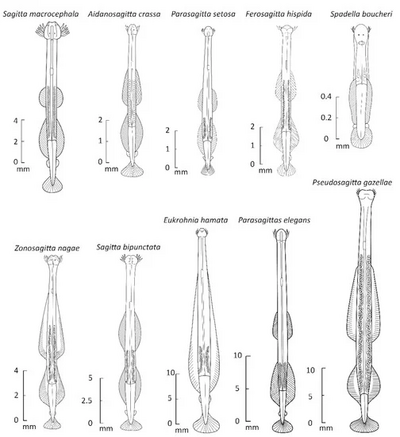
Capinatator praetermissus is an extinct arrow worm from the Burgess Shale lagerstätte, living about 508 million years ago in what is now British Columbia. It has the distinction of having 50 spines around its mouth. As with modern arrow worms, the spines were used to grasp prey for consumption. C. praetermissus is thought to represent a stage of chaetognathan evolution before arrow worms became planktonic swimmers.
Spadellidae is a family of sagittoideans in the order Phragmophora. Spadellidae prey on plankton and commonly reside in the epipelagic zone of the ocean.
Paraspadella is a genus of chaetognaths in the family Spadellidae. Paraspadella was originally considered as Spadella before a revision separated that genus into three genera: Spadella, Paraspadella, and Gephyrospadella, the last of which is now synonymised to Paraspadella. The initial division was based on previous knowledge of three groups of Spadella, in a similar manner in which Sagitta was divided into a family of genera. Paraspadella is differentiated from Spadella by the presence of disparate (digital) adhesive organs, present in the former to various degrees, but entirely absent in the latter.

Timorebestia koprii is an extinct species of stem-group chaetognath that lived about 520 million years ago, in the Cambrian. Its fossils are known from the Sirius Passet Lagerstätte in Greenland, and it was first described in 2024.
Eukrohnia hamata is a species of arrow worm, transparent marine invertebrates belonging to the phylum Chaetognatha. Characterized by its unique morphology and adaptation for life in the deep-sea environment, this planktonic predator is widely distributed throughout the world's oceans, and plays an important role in marine ecosystems.


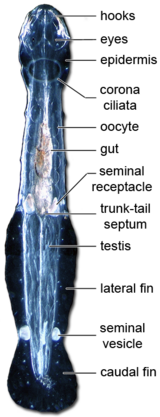



{kind=link}