
A chordate is a deuterostomic animal belonging to the phylum Chordata. All chordates possess, at some point during their larval or adult stages, five distinctive physical characteristics (synapomorphies) that distinguish them from other taxa. These five synapomorphies are a notochord, a hollow dorsal nerve cord, an endostyle or thyroid, pharyngeal slits, and a post-anal tail. The name "chordate" comes from the first of these synapomorphies, the notochord, which plays a significant role in chordate body plan structuring and movements. Chordates are also bilaterally symmetric, have a coelom, possess a closed circulatory system, and exhibit metameric segmentation.

Graptolites are a group of colonial animals, members of the subclass Graptolithina within the class Pterobranchia. These filter-feeding organisms are known chiefly from fossils found from the Middle Cambrian through the Lower Carboniferous (Mississippian). A possible early graptolite, Chaunograptus, is known from the Middle Cambrian. Recent analyses have favored the idea that the living pterobranch Rhabdopleura represents an extant graptolite which diverged from the rest of the group in the Cambrian.

Pterobranchia, members of which are often called pterobranchs, is a class of small worm-shaped animals. They belong to the Hemichordata, and live in secreted tubes on the ocean floor. Pterobranchia feed by filtering plankton out of the water with the help of cilia attached to tentacles. There are about 25 known living pterobranch species in three genera, which are Rhabdopleura, Cephalodiscus, and Atubaria. On the other hand, there are several hundred extinct genera, some of which date from the Cambrian Period.
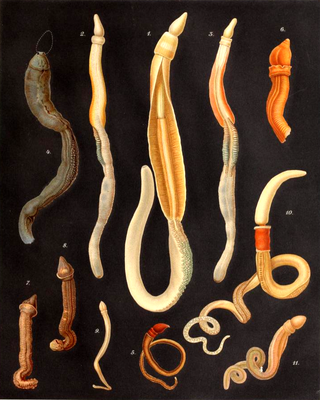
The acorn worms or Enteropneusta are a hemichordate class of invertebrates consisting of one order of the same name. The closest non-hemichordate relatives of the Enteropneusta are the echinoderms. There are 111 known species of acorn worm in the world, the main species for research being Saccoglossus kowalevskii. Two families—Harrimaniidae and Ptychoderidae—separated at least 370 million years ago.

Pharyngeal slits are filter-feeding organs found among deuterostomes. Pharyngeal slits are repeated openings that appear along the pharynx caudal to the mouth. With this position, they allow for the movement of water in the mouth and out the pharyngeal slits. It is postulated that this is how pharyngeal slits first assisted in filter-feeding, and later, with the addition of gills along their walls, aided in respiration of aquatic chordates. These repeated segments are controlled by similar developmental mechanisms. Some hemichordate species can have as many as 200 gill slits. Pharyngeal clefts resembling gill slits are transiently present during the embryonic stages of tetrapod development. The presence of pharyngeal arches and clefts in the neck of the developing human embryo famously led Ernst Haeckel to postulate that "ontogeny recapitulates phylogeny"; this hypothesis, while false, contains elements of truth, as explored by Stephen Jay Gould in Ontogeny and Phylogeny. However, it is now accepted that it is the vertebrate pharyngeal pouches and not the neck slits that are homologous to the pharyngeal slits of invertebrate chordates. Pharyngeal arches, pouches, and clefts are, at some stage of life, found in all chordates. One theory of their origin is the fusion of nephridia which opened both on the outside and the gut, creating openings between the gut and the environment.
In embryology, cleavage is the division of cells in the early development of the embryo, following fertilization. The zygotes of many species undergo rapid cell cycles with no significant overall growth, producing a cluster of cells the same size as the original zygote. The different cells derived from cleavage are called blastomeres and form a compact mass called the morula. Cleavage ends with the formation of the blastula, or of the blastocyst in mammals.

Ambulacraria, or Coelomopora, is a clade of invertebrate phyla that includes echinoderms and hemichordates; a member of this group is called an ambulacrarian. Phylogenetic analysis suggests the echinoderms and hemichordates separated around 533 million years ago. The Ambulacraria are part of the deuterostomes, a clade that also includes the many Chordata, and the few extinct species belonging to the Vetulicolia.

Deuterostomes are bilaterian animals of the superphylum Deuterostomia, typically characterized by their anus forming before the mouth during embryonic development. Deuterostomia is further divided into 4 phyla: Chordata, Echinodermata, Hemichordata, and the extinct Vetulicolia known from Cambrian fossils. The extinct clade Cambroernida is also thought to be a member of Deuterostomia.

Phoronids are a small phylum of marine animals that filter-feed with a lophophore, and build upright tubes of chitin to support and protect their soft bodies. They live in most of the oceans and seas, including the Arctic Ocean but excluding the Antarctic Ocean, and between the intertidal zone and about 400 meters down. Most adult phoronids are 2 cm long and about 1.5 mm wide, although the largest are 50 cm long.

Saccoglossus is a genus of acorn worm. It is the largest genus in this class, with 18 species.
In evolutionary developmental biology, inversion refers to the hypothesis that during the course of animal evolution, the structures along the dorsoventral (DV) axis have taken on an orientation opposite that of the ancestral form.

Harrimaniidae is a basal family of acorn worms. A taxonomic revision was undertaken in 2010, and a number of new genera and species found in the Eastern Pacific were described. In this family the development is direct without tornaria larva, and circular muscle fibers in their trunk is missing. There is some indication that Stereobalanus may be a separate basal acorn worm lineage, sister to all remaining acorn worms.

Rhabdopleura normani is a small, marine species of worm-shaped animal known as a pterobranch. It is a sessile suspension feeder, lives in clear water, and secretes tubes on the ocean floor.
The pterobranchia mitochondrial code is a genetic code used by the mitochondrial genome of Rhabdopleura compacta (Pterobranchia). The Pterobranchia are one of the two groups in the Hemichordata which together with the Echinodermata and Chordata form the three major lineages of deuterostomes. AUA translates to isoleucine in Rhabdopleura as it does in the Echinodermata and Enteropneusta while AUA encodes methionine in the Chordata. The assignment of AGG to lysine is not found elsewhere in deuterostome mitochondria but it occurs in some taxa of Arthropoda. This code shares with many other mitochondrial codes the reassignment of the UGA STOP to tryptophan, and AGG and AGA to an amino acid other than arginine. The initiation codons in Rhabdopleura compacta are ATG and GTG.
Spartobranchus tenuis is an extinct species of acorn worms (Enteropneusta). It existed in the Middle Cambrian. Petrified mark animals were found in British Columbia, Canada in the Burgess Shale formation. It is similar to the modern representatives of the family Harrimaniidae, distinguished by branching fiber tubes. It is a believed predecessor of Pterobranchia, but this species is intermediate between these two classes. Studies show that these tubes were lost in the line leading to modern acorn worm, but remained in the extinct graptolites. However recently Antarctic torquatorids have been found that also make tubes.

Xenambulacraria is a proposed clade of animals with bilateral symmetry as an embryo, consisting of the Xenacoelomorpha and the Ambulacraria.
Billie J. Swalla is a professor of biology at the University of Washington. She was the first female director of Friday Harbor Laboratories, where she worked from 2012 to 2019. Her lab investigates the evolution of chordates by comparative genetic and phylogenetic analysis of animal taxa.

Rhabdopleura compacta is a sessile hemichordate. It is a suspension feeder that secretes tubes on the ocean floor.
Cephalodiscus sibogae is a sessile hemichordate belonging to the order Cephalodiscida. Sightings of the species has been reported only once.

Balanoglossus gigas is a species of large free-living enteropneust found in the Atlantic Ocean. It is the largest acorn worm currently known, and has a strong iodoform-like odour. It is bioluminescent.





