
The Nitrosomonadales are an order of the class Betaproteobacteria in the phylum Pseudomonadota. Like all members of their class, they are Gram-negative.

The Rhodocyclales are an order of the class Betaproteobacteria in the phylum Pseudomonadota ("Proteobacteria"). Following a major reclassification of the class in 2017, the previously monofamilial order was split into three families:

Sphingomonadaceae are a gram-negative bacterial family of the Alphaproteobacteria. An important feature is the presence of sphingolipids in the outer membrane of the cell wall. The cells are ovoid or rod-shaped. Others are also pleomorphic, i.e. the cells change the shape over time. Some species from Sphingomonadaceae family are dominant components of biofilms.
The Hydrogenophilaceae are a family of the class Hydrogenophilalia in the phylum Pseudomonadota ("Proteobacteria"), with two genera – Hydrogenophilus and Tepidiphilus. Like all Pseudomonadota, they are Gram-negative. All known species are thermophilic, growing around 50 °C, and use molecular hydrogen or organic molecules as their source of electrons to support growth; some species are autotrophs.
Thiobacillus is a genus of Gram-negative Betaproteobacteria. Thiobacillus thioparus is the type species of the genus, and the type strain thereof is the StarkeyT strain, isolated by Robert Starkey in the 1930s from a field at Rutgers University in the United States of America. While over 30 "species" have been named in this genus since it was defined by Martinus Beijerinck in 1904,, most names were never validly or effectively published. The remainder were either reclassified into Paracoccus, Starkeya ; Sulfuriferula, Annwoodia, Thiomonas ; Halothiobacillus, Guyparkeria, or Thermithiobacillus or Acidithiobacillus. The very loosely defined "species" Thiobacillus trautweinii was where sulfur oxidising heterotrophs and chemolithoheterotrophs were assigned in the 1910-1960s era, most of which were probably Pseudomonas species. Many species named in this genus were never deposited in service collections and have been lost.

The Acidithiobacillales are an order of bacteria within the class Acidithiobacillia and comprises the genera Acidithiobacillus and Thermithiobacillus. Originally, both were included in the genus Thiobacillus, but they are not related to the type species, which belongs to the Betaproteobacteria.
Acidithiobacillus is a genus of the Acidithiobacillia in the phylum "Pseudomonadota". This genus includes ten species of acidophilic microorganisms capable of sulfur and/or iron oxidation: Acidithiobacillus albertensis, Acidithiobacillus caldus, Acidithiobacillus cuprithermicus, Acidithiobacillus ferrianus, Acidithiobacillus ferridurans, Acidithiobacillus ferriphilus, Acidithiobacillus ferrivorans, Acidithiobacillus ferrooxidans, Acidithiobacillus sulfuriphilus, and Acidithiobacillus thiooxidans.A. ferooxidans is the most widely studied of the genus, but A. caldus and A. thiooxidans are also significant in research. Like all "Pseudomonadota", Acidithiobacillus spp. are Gram-negative and non-spore forming. They also play a significant role in the generation of acid mine drainage; a major global environmental challenge within the mining industry. Some species of Acidithiobacillus are utilized in bioleaching and biomining. A portion of the genes that support the survival of these bacteria in acidic environments are presumed to have been obtained by horizontal gene transfer.

Sulfur-reducing bacteria are microorganisms able to reduce elemental sulfur (S0) to hydrogen sulfide (H2S). These microbes use inorganic sulfur compounds as electron acceptors to sustain several activities such as respiration, conserving energy and growth, in absence of oxygen. The final product of these processes, sulfide, has a considerable influence on the chemistry of the environment and, in addition, is used as electron donor for a large variety of microbial metabolisms. Several types of bacteria and many non-methanogenic archaea can reduce sulfur. Microbial sulfur reduction was already shown in early studies, which highlighted the first proof of S0 reduction in a vibrioid bacterium from mud, with sulfur as electron acceptor and H
2 as electron donor. The first pure cultured species of sulfur-reducing bacteria, Desulfuromonas acetoxidans, was discovered in 1976 and described by Pfennig Norbert and Biebel Hanno as an anaerobic sulfur-reducing and acetate-oxidizing bacterium, not able to reduce sulfate. Only few taxa are true sulfur-reducing bacteria, using sulfur reduction as the only or main catabolic reaction. Normally, they couple this reaction with the oxidation of acetate, succinate or other organic compounds. In general, sulfate-reducing bacteria are able to use both sulfate and elemental sulfur as electron acceptors. Thanks to its abundancy and thermodynamic stability, sulfate is the most studied electron acceptor for anaerobic respiration that involves sulfur compounds. Elemental sulfur, however, is very abundant and important, especially in deep-sea hydrothermal vents, hot springs and other extreme environments, making its isolation more difficult. Some bacteria – such as Proteus, Campylobacter, Pseudomonas and Salmonella – have the ability to reduce sulfur, but can also use oxygen and other terminal electron acceptors.

Campylobacterota are a phylum of Gram-negative bacteria. Only a few genera have been characterized, including the curved to spirilloid Wolinella, Helicobacter, and Campylobacter. Until the 2021 revision of bacterial taxonomy by the ICSP, the entire phylum was classified within the Proteobacteria as the Epsilonproteobacteria.

Betaproteobacteria are a class of Gram-negative bacteria, and one of the eight classes of the phylum Pseudomonadota.

Alphaproteobacteria is a class of bacteria in the phylum Pseudomonadota. The Magnetococcales and Mariprofundales are considered basal or sister to the Alphaproteobacteria. The Alphaproteobacteria are highly diverse and possess few commonalities, but nevertheless share a common ancestor. Like all Proteobacteria, its members are gram-negative, although some of its intracellular parasitic members lack peptidoglycan and are consequently gram variable.
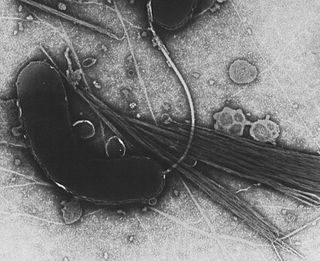
Gammaproteobacteria is a class of bacteria in the phylum Pseudomonadota. It contains about 250 genera, which makes it the most genus-rich taxon of the Prokaryotes. Several medically, ecologically, and scientifically important groups of bacteria belong to this class. All members of this class are Gram-negative. It is the most phylogenetically and physiologically diverse class of the Pseudomonadota.
Thauera is a genus of Gram-negative bacteria in the family Zoogloeaceae of the order Rhodocyclales of the Betaproteobacteria. The genus is named for the German microbiologist Rudolf Thauer. Most species of this genus are motile by flagella and are mostly rod-shaped. The species occur in wet soil and polluted freshwater.
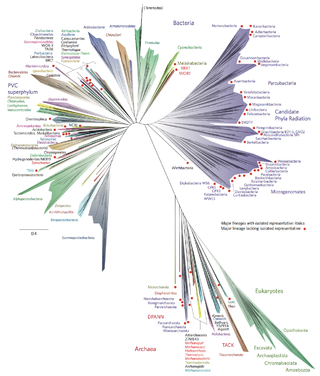
Bacterial phyla constitute the major lineages of the domain Bacteria. While the exact definition of a bacterial phylum is debated, a popular definition is that a bacterial phylum is a monophyletic lineage of bacteria whose 16S rRNA genes share a pairwise sequence identity of ~75% or less with those of the members of other bacterial phyla.

Bacterial taxonomy is subfield of taxonomy devoted to the classification of bacteria specimens into taxonomic ranks.
Dechloromonas is a genus in the phylum Pseudomonadota (Bacteria).
There are several models of the Branching order of bacterial phyla, one of these was proposed in 1987 paper by Carl Woese.
Acidithiobacillia is a class of the phylum Pseudomonadota ("Proteobacteria"). Its type order, the Acidithiobacillales, was formerly classified within the Gammaproteobacteria, and comprises two families of sulfur-oxidising autotrophs, the Acidithiobacillaceae and the Thermithiobacillaceae, which in turn include the genera Acidithiobacillus and Thermithiobacillus.
Ann Patricia Wood is a retired British biochemist and bacteriologist who specialized in the ecology, taxonomy and physiology of sulfur-oxidizing chemolithoautotrophic bacteria and how methylotrophic bacteria play a role in the degradation of odour causing compounds in the human mouth, vagina and skin. The bacterial genus Annwoodia was named to honor her contributions to microbial research in 2017.
