
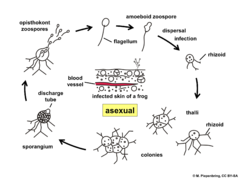
A zoospore is a motile asexual spore that uses a flagellum for locomotion in aqueous or moist environments. Also called a swarm spore, these spores are created by some protists, bacteria, and fungi to propagate themselves. Certain zoospores are infectious and transmittable, such as Batrachochytrium dendrobatidis, a fungal zoospore that causes high rates of mortality in amphibians.
Chytridiomycetes is a class of fungi. Members are found in soil, fresh water, and saline estuaries. They are first known from the Rhynie chert. It has recently been redefined to exclude the taxa Neocallimastigomycota and Monoblepharidomycetes, which are now a phylum and a sister-class respectively.

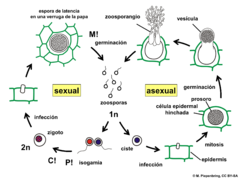
Chytridiomycosis is an infectious disease in amphibians, caused by the chytrid fungi Batrachochytrium dendrobatidis and Batrachochytrium salamandrivorans. Chytridiomycosis has been linked to dramatic population declines or extinctions of amphibian species in western North America, Central America, South America, eastern Australia, east Africa (Tanzania), and Dominica and Montserrat in the Caribbean. Much of the New World is also at risk of the disease arriving within the coming years. The fungus is capable of causing sporadic deaths in some amphibian populations and 100% mortality in others. No effective measure is known for control of the disease in wild populations. Various clinical signs are seen by individuals affected by the disease. A number of options are possible for controlling this disease-causing fungus, though none has proved to be feasible on a large scale. The disease has been proposed as a contributing factor to a global decline in amphibian populations that apparently has affected about 30% of the amphibian species of the world. Some research found evidence insufficient for linking chytrid fungi and chytridiomycosis to global amphibian declines, but more recent research establishes a connection and attributes the spread of the disease to its transmission through international trade routes into native ecosystems.

Batrachochytrium dendrobatidis, also known as Bd or the amphibian chytrid fungus, is a fungus that causes the disease chytridiomycosis in amphibians.
Neocallimastigomycota is a phylum containing anaerobic fungi, which are symbionts found in the digestive tracts of larger herbivores. Anaerobic fungi were originally placed within phylum Chytridiomycota, within Order Neocallimastigales but later raised to phylum level, a decision upheld by later phylogenetic reconstructions. It encompasses only one family.
Hyphochytrids are eukaryotic organisms in the group of Stramenopiles (Heterokonta).

Rhizophydiales are an important group of chytrid fungi. They are found in soil as well as marine and fresh water habitats where they function as parasites and decomposers.
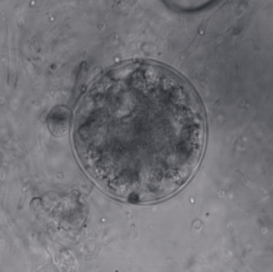
Spizellomycetales is an order of fungi in the Chytridiomycetes. Spizellomycetalean chytrids are essentially ubiquitous zoospore-producing fungi found in soils where they decompose pollen. Recently they have also been found in dung and harsh alpine environments, greatly expanding the range of habitats where one can expect to find these fungi.

Blastocladiomycota is one of the currently recognized phyla within the kingdom Fungi. Blastocladiomycota was originally the order Blastocladiales within the phylum Chytridiomycota until molecular and zoospore ultrastructural characters were used to demonstrate it was not monophyletic with Chytridiomycota. The order was first erected by Petersen for a single genus, Blastocladia, which was originally considered a member of the oomycetes. Accordingly, members of Blastocladiomycota are often referred to colloquially as "chytrids." However, some feel "chytrid" should refer only to members of Chytridiomycota. Thus, members of Blastocladiomycota are commonly called "blastoclads" by mycologists. Alternatively, members of Blastocladiomycota, Chytridiomycota, and Neocallimastigomycota lumped together as the zoosporic true fungi. Blastocladiomycota contains 5 families and approximately 12 genera. This early diverging branch of kingdom Fungi is the first to exhibit alternation of generations. As well, two (once) popular model organisms—Allomyces macrogynus and Blastocladiella emersonii—belong to this phylum.

Entorrhizomycetes is the sole class in the phylum Entorrhizomycota, within the Fungi subkingdom Dikarya along with Basidiomycota and Ascomycota. It contains three genera and is a small group of teliosporic root parasites that form galls on plants in the Juncaceae (rush) and Cyperaceae (sedge) families. Prior to 2015 this phylum was placed under the subdivision Ustilaginomycotina. A 2015 study did a "comprehensive five-gene analyses" of Entorrhiza and concluded that the former class Entorrhizomycetes is possibly either a close sister group to the rest of Dikarya or Basidiomycota.
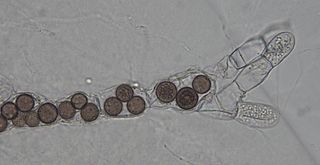
Rozella is a fungal genus of obligate endoparasites of a variety of hosts, including Oomycota, Chytridiomycota, and Blastocladiomycota. Rozella was circumscribed by French mycologist Marie Maxime Cornu in 1872. Considered one of the earliest diverging lineages of fungi, the widespread genus contains 27 species, with the most well studied being Rozella allomycis. Rozella is a member of a large clade of fungi referred to as the Cryptomycota/Rozellomycota. While some can be maintained in dual culture with the host, most have not been cultured, but they have been detected, using molecular techniques, in soil samples, and in freshwater and marine ecosystems. Zoospores have been observed, along with cysts, and the cells of some species are attached to diatoms.

Synchytrium is a large genus of plant pathogens within the phylum Chytridiomycota. Species are commonly known as false rust or wart disease. Approximately 200 species are described, and all are obligate parasites of angiosperms, ferns, or mosses. Early species were mistakenly classified among the higher fungi because of their superficial similarity to the rust fungi. Anton de Bary and Mikhail S. Woronin recognized the true nature of these fungi and established the genus to accommodate Synchytrium taraxaci, which grows on dandelions, and S. succisae, which grows on Succisa pratensis. Synchytrium taraxaci is the type of the genus. The genus has been divided into 6 subgenera based on differences in life cycles.
Olpidium is a fungal genus in the family Olpidiaceae. Members of Olpidium are zoosporic pathogens of plants, animals, fungi, and oomycetes.

Batrachochytrium salamandrivorans (Bsal) is a pathogenic chytrid fungus that infects amphibian species. Although salamanders and newts seem to be the most susceptible, some anuran species are also affected. Bsal has emerged recently and poses a major threat to species in Europe and North America.

Physoderma is a genus of chytrid fungi. Described by German botanist Karl Friedrich Wilhelm Wallroth in 1833, the genus contains some species that are parasitic on vascular plants, including P. alfalfae and P. maydis, causative agents of crown wart of alfalfa and brown spot of corn, respectively. Of the chytrid genera, Physoderma is the oldest. However, species were confused with the rust fungi, the genus Synchytrium, and the genus Protomyces of Ascomycota. Members of Physoderma are obligate parasites of pteridophytes and angiosperms. There are approximately 80 species within this genus.
Neocallimastix patriciarum is a species of fungus that lives in the rumen of sheep and other ruminant species. N. patriciarum is an obligate anaerobe and is an important component of the microbial population within the rumen. Only one of a few rumen fungi, this species is interesting and unique within the fungal world. Originally thought to be a flagellate protists, species within the phylum Neocallimastigomycota were first recognized as a fungus by Colin Orpin in 1975 when he demonstrated that they had cell walls of chitin
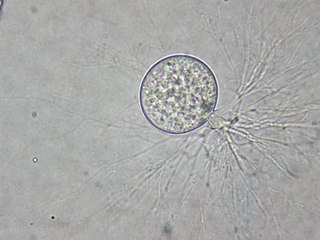
Chytriomyces is the type genus of fungi in the family Chytriomycetaceae. The genus was described by mycologist John Sidney Karling in 1945. The family, created by Peter Letcher in 2011, contains species with a Group I-type zoospore, distinguishing it from Chytridiaceae members, which have a Group II-type zoospore.
Joyce E. Longcore is a mycologist and an associate research professor at the University of Maine. She is most well known for first culturing and describing Batrachochytrium dendrobatidis which is a species of Chytridiomycota fungi that was the first to be known to attack vertebrates. She continues to collect and isolate Chytridiomycota cultures for other researches to use for their own studies.
Spizellomyces punctatus is a chytrid fungus living in soil. It is a saprotrophic fungus that colonizes decaying plant material. Being an early diverging fungus, S. punctatus retains ancestral cellular features that are also found in animals and amoebae. Its pathogenic relatives, Batrachochytrium dendrobatidis and B. salamandrivorans, infect amphibians and cause global biodiversity loss. The pure culture of S. punctatus was first obtained by Koch.
Marilyn Rose Noyes Mollicone was an American botanist advancing mycology in Maine and advocating for naturalist education.





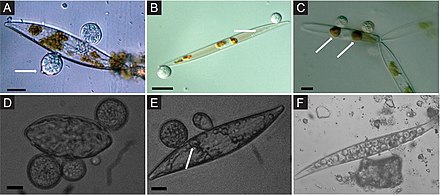
![Pennate diatom from an Arctic meltpond, infected with two chytrid-like [zoo-]sporangium fungal pathogens (in false-colour red). Pennate diatom infected with two chytrid-like fungal pathogens.png](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b5/Pennate_diatom_infected_with_two_chytrid-like_fungal_pathogens.png/440px-Pennate_diatom_infected_with_two_chytrid-like_fungal_pathogens.png)
